Temporal Binding Windows: A Novel Framework for Episodic Memory Research and Psychotherapeutic Drug Development
This article synthesizes cutting-edge research on temporal binding tasks as a paradigm for investigating episodic memory.
Temporal Binding Windows: A Novel Framework for Episodic Memory Research and Psychotherapeutic Drug Development
Abstract
This article synthesizes cutting-edge research on temporal binding tasks as a paradigm for investigating episodic memory. Targeting researchers and drug development professionals, it explores the fundamental neural mechanisms of how the brain binds temporal features into coherent episodic memories. The scope extends from foundational concepts and methodological applications in psychopharmacology to troubleshooting experimental challenges and validating findings through cross-population and computational modeling approaches. Special emphasis is placed on how psychoactive drugs modulate emotional episodic memory encoding, consolidation, and retrieval, offering crucial insights for developing novel therapeutic interventions for addiction, affective disorders, and neurodegenerative conditions.
The Neural Architecture of Temporal Binding in Episodic Memory Formation
Defining Temporal Binding Windows (TBW) and Their Role in Episodic Coherence
The Temporal Binding Window (TBW) is a fundamental concept in neuroscience that describes the epoch of time within which stimuli from different sensory modalities are highly likely to be integrated and perceptually bound into a unified event [1]. This temporal window represents a critical period for multisensory integration, allowing the brain to determine whether sensory inputs originating from different senses (e.g., auditory and visual) belong to the same environmental event [2] [3]. The construct of the TBW has gained significant importance in understanding how the brain constructs coherent perceptual experiences from continuous sensory inputs and how these processes relate to episodic memory formation and coherence [4] [5].
The brain faces considerable challenges in coordinating sensory information due to differences in the physical propagation speeds of environmental energies and variations in neural processing times across sensory modalities [3]. For example, visual stimuli typically require longer processing times compared to auditory stimuli [3]. The TBW provides a temporal solution to this problem by defining a period of tolerance for stimulus asynchrony, thus enabling the binding of multisensory information that likely arises from a common source [1]. The width of the TBW reflects the temporal resolution of this integration system, with narrower windows indicating more precise temporal processing [2].
Table 1: Key Characteristics of Temporal Binding Windows
| Characteristic | Description | Functional Significance |
|---|---|---|
| Window Width | Temporal span for multisensory integration | Narrower windows indicate more precise temporal processing [2] |
| Stimulus Specificity | Varies by stimulus complexity | Smallest for simple flashes/beeps, largest for complex speech [1] |
| Point of Subjective Simultaneity (PSS) | Temporal offset for perceived simultaneity | Typically occurs with auditory lag due to neural processing differences [1] |
| Developmental Trajectory | Matures throughout adolescence | Broad in childhood, narrows through adolescence into adulthood [1] |
| Neural Basis | Supported by multisensory cortical and subcortical structures | Includes superior colliculus, auditory cortex, and association areas [1] |
Neural Mechanisms and Computational Principles
The neural architecture supporting temporal binding involves a distributed network of cortical and subcortical structures where information from multiple senses converges [1]. Neurophysiological studies have revealed that multisensory neurons in areas such as the superior colliculus exhibit temporal tuning functions that underlie the behavioral manifestations of the TBW [1]. These neurons respond most strongly to multisensory stimuli that occur in close temporal proximity, with response enhancement declining as the temporal disparity between stimuli increases [1].
The TBW operates according to several key principles that optimize multisensory integration. The temporal principle dictates that multisensory stimuli occurring in close temporal proximity are more likely to be integrated [1]. This principle is complemented by the spatial principle (favoring stimuli from proximate locations) and the principle of inverse effectiveness (greater integration benefits for weaker stimuli) [1]. Recent research suggests these principles are interdependent rather than operating in isolation, with the brain dynamically weighting their influence based on task demands and context [1].
The computational processes underlying temporal binding involve the detection of temporal coherence across sensory channels [6]. Neural populations encoding correlated sensory features tend to synchronize their activity, facilitating the binding of these features into coherent perceptual representations [6]. This temporal coherence detection provides a mechanism for segmenting continuous sensory input into discrete perceptual events, which serves as the foundation for episodic memory formation [5] [6].
Experimental Assessment of Temporal Binding Windows
Psychophysical Paradigms and Protocols
Researchers employ well-established psychophysical paradigms to quantify the TBW in human participants. Two primary tasks used for this assessment are the Simultaneity Judgment (SJ) task and the Temporal Order Judgment (TOJ) task [3]. In both paradigms, participants are typically seated in a dark, quiet room approximately 48-60 cm from a visual display with auditory stimuli presented via headphones [2] [3].
Simultaneity Judgment (SJ) Task Protocol:
- Stimuli: Pairs of visual and auditory stimuli (e.g., white ring visual stimulus and 1800 Hz tone burst) presented at varying stimulus onset asynchronies (SOAs) [2]
- SOA Range: Typically from -300 ms (auditory leading) to +300 ms (visual leading) in 50 ms steps [2]
- Trial Structure: Fixation crosshair appears 1000 ms before stimulus presentation; stimuli presented briefly (visual: 8.3 ms, auditory: 10 ms) [2]
- Task: Participants indicate whether stimuli were "simultaneous" or "non-simultaneous" via button press [2]
- Data Analysis: Proportion of "simultaneous" responses plotted as a function of SOA; TBW defined as range of SOAs yielding simultaneity responses above a specific threshold (e.g., 75%) [3]
Temporal Order Judgment (TOJ) Task Protocol:
- Stimuli: Similar to SJ task but participants indicate which modality appeared first [3]
- Task: Participants judge temporal order (e.g., "visual first" vs. "auditory first") [3]
- Data Analysis: Psychometric functions fitted to identify SOA where order discrimination is at chance level (PSS) and range where performance is above chance (TBW width) [3]
Table 2: Standard Stimulus Parameters for TBW Assessment
| Parameter | Specification | Typical Values |
|---|---|---|
| Visual Stimulus | White ring on black background | 15° visual angle, 8.3 ms duration [2] |
| Auditory Stimulus | Pure tone burst | 1800 Hz, 10 ms duration, 110 dB SPL [2] |
| SOA Range | Interval between stimulus onsets | -300 to +300 ms in 50 ms steps [2] |
| Trial Number | Assessments per participant | 325 trials for full assessment [2] |
| Response Modality | Button press | 2-alternative forced choice [2] |
Data Analysis and Parameter Extraction
From SJ and TOJ tasks, researchers extract two primary parameters: the Temporal Binding Window width and the Point of Subjective Simultaneity (PSS). The TBW width represents the range of SOAs within which participants perceive simultaneity, typically operationalized as the SOA range where simultaneity responses exceed a specific threshold [3]. The PSS represents the specific SOA at which participants are most likely to perceive the stimuli as simultaneous, which typically shows a slight auditory lag due to differences in neural processing times between modalities [1] [3].
The TBW is typically modeled using Gaussian or sigmoidal functions fitted to the proportion of simultaneity responses across SOAs. The window width is often defined as the range between the SOAs corresponding to 25% and 75% of the maximum simultaneity response rate [3]. More recent approaches use cumulative normal distributions to model the data, with the window width corresponding to the standard deviation of the distribution [2].
Temporal Binding and Episodic Coherence
Role in Event Segmentation and Memory Formation
Temporal binding processes play a crucial role in segmenting continuous experience into discrete events, which forms the foundation of episodic memory [5]. The brain employs temporal integration across multiple timescales, from sub-second sensory processing to longer-term event integration, to create coherent episodic representations [5]. This segmentation process allows for the organization of experience into meaningful units that can be efficiently encoded and retrieved from memory.
Recent research has revealed that negative emotion reduces the temporal compression of events in episodic memory, suggesting that emotional valence influences how temporal information is bound into memory representations [4]. In experiments where participants mentally replayed videos depicting negative or neutral events, the time taken to mentally replay negative videos was significantly longer relative to their actual duration compared to neutral videos [4]. This indicates that negative emotion increases the sampling rate of experiential units, leading to lower compression of events in memory representations [4].
Neural Substrates Linking TBW and Episodic Memory
The neural circuits supporting temporal binding show significant overlap with those underlying episodic memory formation. Multisensory integration areas in the temporal and parietal cortices interact with medial temporal lobe structures, particularly the hippocampus, to bind sensory features with temporal context information [5]. This interaction supports the creation of coherent episodic memories that integrate what, where, and when information about experienced events.
The precision of temporal binding directly influences the quality and coherence of episodic memories. Narrower TBWs are associated with more precise temporal encoding of event features, leading to more accurate and detailed memory representations [4] [5]. Conversely, widened TBWs result in less precise binding of temporal information, potentially contributing to fragmented or less coherent memory representations [3].
Clinical Implications and Alterations in Disorders
TBW Alterations in Neurodevelopmental and Neurodegenerative Conditions
Substantial evidence demonstrates that the TBW is altered in various neurodevelopmental and neurodegenerative disorders. Individuals with autism, dyslexia, and schizophrenia consistently show widened TBWs, indicating less precise temporal processing of multisensory information [1]. These alterations in temporal binding may contribute to the perceptual and cognitive difficulties characteristic of these conditions [1].
In neurodegenerative conditions, research has shown that patients with mild cognitive impairment (MCI) and Alzheimer's disease (AD) exhibit extended TBWs compared to healthy older adults [3]. This temporal processing deficit may contribute to the cognitive disorientation and memory fragmentation experienced by these patients. The TBW has been proposed as a potential sensitive marker for early detection and monitoring of cognitive decline [3].
TBW Alterations Following Acquired Brain Injury
Recent research has demonstrated that individuals with a history of concussion show significant widening of the TBW, particularly in temporal order judgment tasks [3]. This impairment persists even after other symptoms have resolved and shows a negative correlation with time since injury, suggesting gradual recovery of temporal processing precision [3]. These findings indicate that TBW assessment may provide a sensitive measure for guiding return-to-activity decisions following concussion [3].
Figure 1: Impact of TBW Alterations on Episodic Memory in Clinical Populations
Table 3: TBW Alterations in Clinical Populations
| Clinical Condition | TBW Alteration | Functional Consequences |
|---|---|---|
| Autism Spectrum Disorder | Widened TBW [1] | Impaired multisensory integration, sensory overload [1] |
| Schizophrenia | Widened TBW [1] | Perceptual fragmentation, reality monitoring deficits [1] |
| Dyslexia | Widened TBW [1] | Auditory-visual integration deficits affecting reading [1] |
| Alzheimer's Disease/MCI | Widened TBW [3] | Temporal disorientation, memory fragmentation [3] |
| Post-Concussion Syndrome | Widened TBW [3] | Impaired temporal order judgment, delayed recovery [3] |
Plasticity and Training of Temporal Binding
Perceptual Training Interventions
Research has demonstrated that the TBW is highly plastic and can be modified through targeted perceptual training. In a seminal study, participants underwent perceptual training using a simultaneity judgment task with feedback, resulting in a approximately 40% narrowing of the TBW after just five hours of training [2]. This training effect was observed using both two-alternative forced choice (2-AFC) and two-interval forced choice (2-IFC) paradigms, indicating robust plasticity in multisensory temporal processing [2].
The training protocol involved:
- Pretraining Assessment: Baseline SJ task across 13 SOAs from -300 to +300 ms [2]
- Training Blocks: 3 blocks per day for 5 days using only SOAs between -150 and +150 ms with immediate feedback [2]
- Feedback: "Correct!" with happy face or "Incorrect" with sad face presented for 500 ms after each response [2]
- Post-training Assessment: Identical to pretraining assessment to measure training effects [2]
- Follow-up: Assessment after one week without training to evaluate retention [2]
This perceptual learning paradigm demonstrates that the neural circuits underlying temporal binding remain malleable into adulthood, with potential implications for therapeutic interventions in clinical populations with widened TBWs [2].
Mechanisms of Training-Induced Plasticity
The neural mechanisms underlying training-induced narrowing of the TBW likely involve enhanced temporal precision in multisensory neural populations and improved top-down regulation of sensory integration processes [2]. Perceptual training may strengthen inhibitory circuits that sharpen temporal tuning, resulting in more selective integration of simultaneous stimuli and reduced binding of asynchronous stimuli [2].
Training effects appear to reflect genuine changes in perceptual sensitivity rather than response bias shifts, as demonstrated by transfer of training effects across different task paradigms [2]. The retention of training effects after one week suggests that these interventions can produce relatively lasting modifications to temporal processing mechanisms [2].
Research Toolkit: Methods and Reagents
Essential Research Equipment and Software
Table 4: Essential Research Equipment for TBW Studies
| Equipment/Software | Specification | Research Application |
|---|---|---|
| Visual Display | High refresh-rate monitor (120 Hz) [2] | Precise visual stimulus timing |
| Auditory System | Calibrated headphones with sound level meter [2] | Controlled auditory presentation |
| Response Collection | Response box (e.g., Psychology Software Tools) [2] | Accurate response time measurement |
| Experimental Control | E-Prime, PsychoPy, or MATLAB with Psychtoolbox | Stimulus presentation and data collection |
| Timing Verification | Oscilloscope with <10 ms error tolerance [2] | Validation of stimulus timing accuracy |
Standardized Stimulus Parameters
For rigorous assessment of temporal binding, researchers should adhere to standardized stimulus parameters:
- Visual Stimuli: Simple geometric forms (circles, rings) with high luminance contrast, brief durations (5-20 ms), subtending approximately 10-15° of visual angle [2]
- Auditory Stimuli: Pure tones (1000-2000 Hz) of brief duration (10-50 ms) at clearly audible but comfortable levels (70-80 dB SPL) [2] [3]
- Spatial Arrangement: Stimuli typically presented in central fixation to control for spatial attention effects [2]
- Experimental Environment: Dark, quiet room to minimize extraneous sensory stimulation [2] [3]
Future Directions and Research Applications
Integration with Episodic Memory Research
Future research should further elucidate the specific mechanisms linking temporal binding processes to episodic memory formation and coherence. Promising directions include examining how individual differences in TBW width predict the temporal precision of episodic memories and investigating how temporal binding deficits contribute to memory impairments in clinical populations [4] [5].
The relationship between temporal binding and emotion warrants further exploration, particularly given recent findings that negative emotion reduces temporal compression in memory [4]. Understanding how emotional valence influences the sampling rate of experiential units could provide important insights into emotional memory formation and trauma-related memory disturbances.
Therapeutic Applications and Interventions
The demonstrated plasticity of the TBW suggests potential therapeutic applications for populations with widened temporal windows [2]. Targeted perceptual training could be developed as an intervention to improve multisensory integration and temporal processing in disorders such as autism, schizophrenia, and post-concussion syndrome [1] [2] [3].
Future research should explore whether TBW narrowing through training produces functional benefits in everyday activities requiring precise temporal processing, such as speech perception in noisy environments, motor coordination, and social interaction. Longitudinal studies are needed to determine the durability of training effects and optimal protocols for maintaining benefits over time [2].
Figure 2: Future Research Directions for TBW and Episodic Coherence
In conclusion, the Temporal Binding Window represents a crucial interface between sensory processing and higher cognitive functions, particularly episodic memory. Its assessment provides valuable insights into the temporal architecture of perception and memory, with broad implications for understanding both typical and atypical cognitive functioning. The continued investigation of TBW and its role in episodic coherence promises to advance both theoretical models and clinical applications in cognitive neuroscience.
Episodic memory, the ability to recall the "what," "where," and "when" of past experiences, requires the binding of disparate event elements into unified representations. The medial temporal lobe (MTL), particularly the hippocampus, serves as the central hub for this binding process, working in concert with a distributed cortical network [7]. Object-location memory represents a fundamental aspect of this system, requiring the formation of associations between specific items and their spatial contexts. Contemporary research has revealed that the hippocampus does not function in isolation but rather participates in dynamic hippocampal-cortical interactions that support both the initial formation and long-term consolidation of these associative memories [8] [9]. This application note details the experimental protocols and analytical frameworks for investigating these neurobiological substrates, providing researchers with standardized methodologies for probing the neural mechanisms of object-location binding within the context of temporal binding tasks for episodic memory research.
Core Neural Mechanisms of Object-Location Binding
Hippocampal Role in High-Resolution Associative Binding
The hippocampus is specialized for generating complex high-resolution bindings that link qualitative aspects of an event [10]. This binding function supports recollection, the retrieval of qualitative information about a prior event, which can be contrasted with familiarity-based recognition that lacks specific contextual details. Patient studies have established that hippocampal damage leads to selective deficits in recollection while largely sparing familiarity-based recognition [10].
Table 1: Neural Coding Strategies in the Hippocampal System
| Brain Region | Coding Strategy | Spatial Resolution | Representational Properties |
|---|---|---|---|
| Dorsal Hippocampus | Sparse coding | High local precision | Smaller place fields; region-specific information |
| Ventral Hippocampus | Dense coding | Broad spatial distribution | Informative over larger environmental extents |
| Medial Entorhinal Cortex | Grid cell tessellation | Metric spatial framework | Repetitive hexagonal firing patterns |
Recent single-neuron recordings in humans have identified episode-specific neurons (ESNs) in the hippocampus that fire in response to the conjunction of all elements within a specific episode rather than individual content elements [11]. These ESNs, observed exclusively in the hippocampus, code discrete episodic memories using either rate codes or temporal firing codes, and do not merely respond to invariant elements or specific temporal windows [11].
Cortical and Hippocampal-Cortical Contributions
The prefrontal cortex (PFC), particularly the infralimbic (IL) cortex, plays a critical role in storing consolidated social memories [8]. Inactivation of IL neurons projecting to the nucleus accumbens shell (IL→NAcSh) impairs social recognition, demonstrating the essential role of cortical substrates in maintaining familiar representations [8].
The complementary learning systems framework posits that the hippocampus serves as a fast-learner for specific episodes, while the neocortex serves as a slow-learner that extracts generalized knowledge across experiences [12]. Recent experimental evidence confirms that naturally restricted plasticity in the neocortex protects previous knowledge from interference, while artificially increasing cortical plasticity leads to better one-trial memory but increased interference in semantic-like memory [12].
Experimental Protocols for Investigating Object-Location Binding
Multivoxel Pattern Analysis (MVPA) of Sequence Memory
Objective: To characterize hippocampal coding of object and temporal context information during retrieval of learned object sequences [13].
Participants: Human subjects with normal neurological profiles.
Apparatus: Functional MRI scanner with standard head coil; presentation software for visual stimuli.
Stimuli: Multiple object sequences including:
- Fixed sequence: Five objects not overlapping with other sequences
- Overlapping sequences (X1/X2): Shared objects in positions 2 and 3
- Overlapping sequences (Y1/Y2): Shared objects in the first three positions
- Random sequence: Same five objects presented in random order
Procedure:
- Learning Session (Pre-scan):
- Participants learn five object sequences by making semantic decisions about each object
- Training continues until performance criteria are met (e.g., >90% accuracy)
fMRI Scanning Session:
- Participants make semantic decisions on a continuous stream of objects
- Stimulus presentation: Contiguous presentations of the five learned sequences and one random sequence
- No obvious boundaries between sequences during scanning
- fMRI parameters: Standard whole-brain coverage with high-resolution structural and functional sequences
Behavioral Measures:
- Accuracy and reaction times for semantic judgments
- Analysis of sequence position effects on performance
Multivoxel Pattern Analysis:
- Preprocessing: Standard fMRI preprocessing pipeline (motion correction, normalization, etc.)
- Region of Interest (ROI) definition: Hippocampus, parahippocampal cortex, perirhinal cortex
- Pattern similarity analysis: Calculate correlations in voxel activity patterns between pairs of trials that share specific information
- Statistical testing: Compare pattern similarity across conditions of interest
Analysis Focus:
- Information about temporal positions of objects in learned vs. random sequences
- Differentiation between overlapping object sequences
- Differentiation between temporally adjacent objects from distinct sequence contexts
- Comparison of pattern information profiles across hippocampal subregions and cortical areas
Single-Neuron Recording of Episode-Specific Coding
Objective: To identify episode-specific neurons (ESNs) in the human hippocampus that code for discrete episodic memories [11].
Participants: Patients with medically intractable epilepsy implanted with stereotactic depth electrodes for clinical monitoring.
Apparatus: Intracranial microwire recordings from hippocampus and parahippocampal regions; stimulus presentation system.
Stimuli: Unique image associations for each episode (animals, faces, places).
Procedure:
- Encoding Phase:
- Participants create vivid mental stories consisting of an animal cue and associated images
- Self-paced presentation with minimum viewing duration enforced
Distractor Task:
- Odd/even number judgments (22.43-224.52 second duration)
- Prevents maintenance rehearsal
Recall Phase:
- Animal cue presentation with request to retrieve associated images
- Behavioral measure of recall accuracy
Visual Tuning Task (Experiment 2):
- Repeated presentation of images from memory task
- Identifies concept neurons responsive to specific invariant elements
Neuronal Recording:
- Continuous recording during all task phases
- Spike sorting and isolation of single units
Analysis Pipeline:
- Firing Rate Calculation:
- Determine firing rate during each episode at encoding and retrieval
- Z-score firing rates across all encoding and retrieval episodes
- Exclude forgotten episodes
ESN Identification:
- Measure episode-specific firing reinstatement as product of standardized firing rates at encoding and retrieval
- Generate null distribution using episode-shuffling procedure
- Classify ESNs using dual criteria:
- Empirical reinstatement value >99th percentile of shuffled distribution for at least one episode
- Standardized firing rate for encoding and retrieval each >1.645 (p<0.05, one-tailed)
Control Analyses:
- Exclude neurons with significant firing increases during first second after animal cue encoding
- Test for concept neuron properties using visual tuning task data
- Verify specificity to hippocampus vs. parahippocampal regions
Table 2: Key Research Reagent Solutions for Hippocampal-Cortical Network Research
| Reagent/Technique | Function/Application | Key Characteristics | Experimental Utility |
|---|---|---|---|
| RGS14414 Gene | Plasticity enhancer via viral overexpression | Increases BDNF and dendritic branching in targeted areas | Testing cortical plasticity thresholds in memory interference paradigms [12] |
| Halorhodopsin (NpHR) | Optogenetic inhibition | Light-activated neural silencing | Temporally-precise inactivation of specific neural populations during memory phases [8] |
| Designer Receptors Exclusively Activated by Designer Drugs (DREADDs) | Chemogenetic manipulation | Chemically-activated neural modulation | Longer-term manipulation of neural activity without implanted hardware [8] |
| GCaMP6f Calcium Indicator | In vivo calcium imaging | Genetically-encoded calcium sensor | Monitoring population-level neural activity in behaving animals [8] |
| Multivoxel Pattern Analysis (MVPA) | fMRI pattern classification | Multivariate analysis of distributed activity patterns | Decoding cognitive states and information content from BOLD signals [13] |
Visualization of Hippocampal-Cortical Circuitry and Workflows
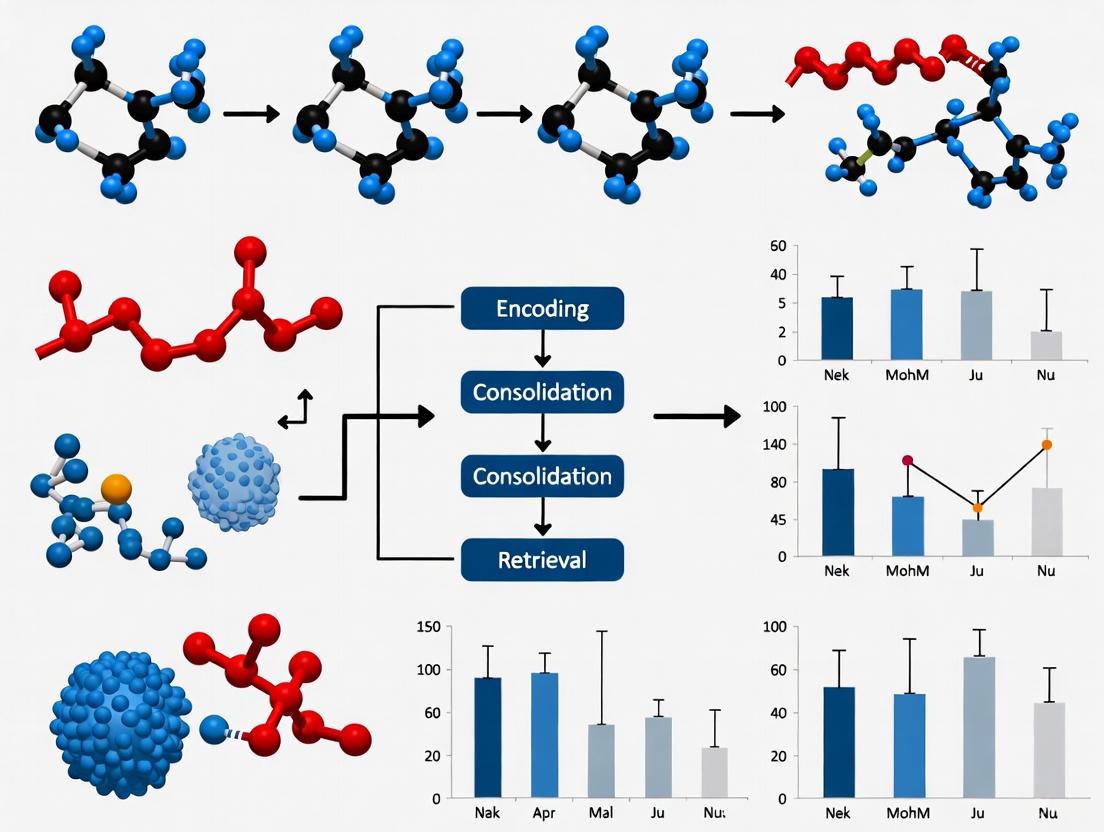
Diagram 1: Hippocampal-Cortical Circuitry for Object-Location Memory. This diagram illustrates the major information pathways supporting object-location binding, highlighting the convergence of object identity (via perirhinal cortex) and spatial context (via parahippocampal cortex) in the hippocampus, with downstream interactions with prefrontal cortex supporting cognitive control and consolidation.
Diagram 2: Temporal Binding Task Experimental Workflow. This diagram outlines the sequential phases of a standardized object-location binding experiment, highlighting critical cognitive processes and corresponding neural engagement across encoding, consolidation, and retrieval phases.
Quantitative Data Synthesis
Table 3: Behavioral and Neural Correlates in Object-Location Binding Tasks
| Experimental Measure | Finding | Experimental Context | Neural Correlate |
|---|---|---|---|
| Reaction Time Advantage | Significantly faster RTs for learned sequences vs. random sequences (F₁,₁₈=36.018, p<0.001) [13] | Object sequence retrieval with semantic decisions | Hippocampal pattern separation of temporal context |
| Episode-Specific Neurons | 23.25% of hippocampal neurons (136/585) classified as ESNs (p<0.001) [11] | Human single-unit recording during associative memory | Hippocampal conjunctive coding of episodic elements |
| Memory Interference | Increased cortical plasticity enhanced one-trial memory but impaired semantic-like memory (p<0.01) [12] | Object Space Task with RGS14414 cortical overexpression | Cortical learning rate modulation of interference susceptibility |
| Social Memory Specificity | Inactivation of IL→NAcSh neurons impaired familiar conspecific recognition [8] | Social familiarization/recognition task in mice | Cortical storage of consolidated social memories |
| Sparse vs. Dense Coding | Equivalent spatial decoding accuracy from dorsal (sparse) and ventral (dense) hippocampal ensembles [9] | Rat open field foraging with population analysis | Complementary coding strategies supporting similar information |
Application in Drug Development and Translational Research
The protocols and mechanisms described herein provide valuable frameworks for evaluating cognitive-enhancing therapeutics and investigating pathological memory processes. Key applications include:
Target Validation: Hippocampal-cortical interaction metrics serve as biomarkers for target engagement in cognitive disorders. The identification of specific binding deficits can guide therapeutic development for conditions like Alzheimer's disease where object-location memory impairment represents an early symptom.
Cognitive Endophenotyping: Standardized object-location binding tasks enable stratification of patient populations based on specific mnemonic deficits (e.g., recollection vs. familiarity impairments), facilitating targeted clinical trials.
Mechanism of Action Studies: The experimental protocols allow precise dissection of whether cognitive enhancers act primarily on hippocampal binding processes, cortical consolidation mechanisms, or hippocampal-cortical interactions.
Interference Management: Understanding how cortical plasticity thresholds protect against memory interference [12] informs development of treatments that balance memory enhancement with stability of existing knowledge.
These applications underscore the translational value of precisely characterizing hippocampal-cortical networks in object-location binding, providing a neurobiologically-grounded framework for developing interventions targeting the core cognitive architecture of episodic memory.
Application Notes
This document provides application notes and experimental protocols for investigating the segmental structure of events and its role in temporal memory compression. This research area integrates principles from episodic memory and temporal binding to understand how continuous experience is segmented into units and how the temporal relationships between these units are distorted in memory. The content is framed within the context of employing temporal binding tasks for advanced episodic memory research, particularly relevant for developing cognitive biomarkers in neurodegenerative and neuropsychiatric drug development.
Table 1: Key Findings on Segmental Structure and Temporal Memory Compression
| Key Finding | Experimental Paradigm | Measured Outcome | Relevance to Temporal Binding & Episodic Memory |
|---|---|---|---|
| Working Memory (WM) modulates temporal compression [14] | Dual-task paradigm during video viewing (High vs. Low Event Boundaries) | Temporal compression rate (mental replay time vs. actual time); Number of recalled experience units [14] | WM availability during encoding shapes the fidelity of event segments, directly impacting the temporal structure of the resulting memory trace. |
| Semantic structure guides recall across age groups [15] | Naturalistic video encoding with multiple recalls over a week; Network analysis of narrative semantic similarity [15] | Recall of central (gist) vs. peripheral details; Influence of semantic connectivity on recall probability [15] | The semantic network of an event influences memory reconstruction, a process that can be implicit in temporal binding of causally related elements. |
| Temporal binding reduces perceptual uncertainty [16] | Interval estimation with masked visual markers | Perceived duration of intervals between markers; Estimation thresholds (precision) [16] | Binding of interval markers serves a functional role in minimizing variance in subjective duration, a mechanism that may extend to binding of event segments in memory. |
| Action cues influence temporal binding [17] | Interval estimation between self-initiated actions and their effects with congruent/incongruent verbal cues | Compression of perceived interval between action and effect (intentional binding) [17] | Demonstrates how inferential (social) cues can modulate the implicit sense of agency and temporal perception, linking cognitive context to temporal binding. |
The core principle is that continuous experience is automatically segmented into discrete experience units at event boundaries, which are meaningful shifts in time, place, or context [15]. These units are the building blocks of episodic memory. Temporal compression—the phenomenon where the mental replay of an event is shorter than its actual duration—is a key metric for understanding how these segments are stored and retrieved [14]. Research indicates that both the inherent segmental structure of an experience and the availability of cognitive resources like working memory during encoding are critical factors shaping this compression [14].
Furthermore, the semantic relatedness between event segments facilitates their recall, a effect preserved in aging, though the recall of peripheral details diminishes [15]. This aligns with the temporal binding phenomenon, where related events or actions and their outcomes are perceived as closer in time than they actually are [17]. This binding is thought to be a fundamental mechanism for constructing coherent episodic narratives, reducing perceptual and memory uncertainty by linking related elements [16].
Experimental Protocols
Protocol 1: Dual-Task Paradigm for Assessing Working Memory's Role in Temporal Compression
This protocol is adapted from Leroy et al. to investigate how limited WM resources during encoding affect the temporal structure of episodic memory [14].
1. Objectives:
- To determine the effect of a concurrent WM load on the number of experience units encoded from a narrative.
- To measure the impact of WM load on the temporal compression rate during mental replay.
2. Materials:
- Stimuli: A set of 1-minute videos pre-analyzed for a high or low number of event boundaries [14].
- WM Task: A secondary, continuous task such as an auditory n-back task or a visual pattern tracking task.
- Data Collection Software: Experiment builder (e.g., Gorilla SC[https://gorilla.sc]) for precise stimulus presentation and response collection [15].
3. Procedure:
- Participant Preparation: Obtain informed consent. Provide standardized instructions.
- Encoding Phase (Within-Subjects Design):
- Condition A (No Load): Participant watches a video and is instructed to remember it for a later test.
- Condition B (WM Load): Participant watches a different video while simultaneously performing the concurrent WM task.
- Video types (high/low boundaries) and conditions are counterbalanced.
- Immediate Test Phase (after each video):
- Temporal Replay Task: Participant is instructed to mentally replay the video event in their mind "as accurately as possible" and to press a key when finished. The mental replay time is recorded.
- Verbal Recall: Participant provides a verbal description of everything they can remember from the video. Responses are audio-recorded.
- Data Analysis:
- Temporal Compression Rate: Calculate as
(1 - (Mental Replay Time / Actual Video Duration)) * 100[14]. - Experience Units Recalled: Transcribe verbal recalls. The number of discrete experience units (distinct actions, sub-events) is coded by trained raters or using NLP tools.
- Temporal Compression Rate: Calculate as
Figure 1: Experimental workflow for the dual-task paradigm, assessing working memory's role in temporal compression.
Protocol 2: Naturalistic Event Recall to Probe Semantic Structure and Gist-Based Memory
This protocol is adapted from the study on semantic structure in young and older adults, ideal for testing populations where cognitive aging or neurodegeneration is a factor [15].
1. Objectives:
- To quantify the influence of semantic similarity between event units on their subsequent recall.
- To compare the recall of central (gist) versus peripheral details across multiple time delays and age groups.
2. Materials:
- Stimuli: 8 short videos (approx. 3-4 minutes each) depicting life situations with a clear narrative [15].
- Recall Platform: Online testing platform (e.g., Gorilla) or lab-based setup with audio recording capabilities [15].
3. Procedure:
- Session 1 (Day 1 - Encoding & Immediate Recall):
- Encoding: Participant watches all 8 videos in a pseudo-randomized order. Each video is preceded by a title.
- Immediate Recall: Participant is cued with the titles of 4 randomly selected videos and asked to verbally describe each in as much detail as possible.
- Session 2 (Day 2 - 24-hour Delay Recall): Participant is again cued to recall the same 4 videos from Day 1.
- Session 3 (Day 8 - 1-week Delay Recall): Participant is cued to recall all 8 original videos.
- Data Analysis:
- Narrative Transcription: Verbal recalls are transcribed verbatim.
- Detail Classification: Transcribed details are coded as:
- Central Details: Essential to the storyline and its overall meaning.
- Peripheral Details: Contextual and perceptual information that enriches the narrative [15].
- Semantic Network Analysis: Narrative descriptions for each video are transformed into a network where nodes are events and edges represent their semantic similarity. Centrality metrics (e.g., degree centrality) are computed for each event [15].
Figure 2: Multi-session protocol for probing semantic structure and gist-based memory over time.
The Scientist's Toolkit
Table 2: Essential Research Reagents and Materials
| Item | Function/Description | Example Application in Protocol |
|---|---|---|
| Naturalistic Video Stimuli | Short films depicting life situations with a clear narrative structure. Essential for ecological validity [15]. | Core stimulus for Probing Semantic Structure (Protocol 2). Videos are pre-analyzed for event boundaries. |
| Event Boundary Annotations | Pre-defined segmentation of videos into discrete experience units based on shifts in time, place, or context [15] [14]. | Provides the ground truth for analyzing recalled experience units in both protocols. |
| Dual-Task Load | A secondary cognitive task (e.g., n-back, pattern tracking) designed to consume working memory resources during encoding [14]. | Critical manipulation in the Dual-Task Paradigm (Protocol 1) to test resource dependence of segmentation. |
| Temporal Replay Task | A behavioral measure where participants mentally replay an event; the duration indicates temporal compression [14]. | Key dependent variable in Protocol 1. Mental replay time is compared to actual event duration. |
| Central/Peripheral Detail Coding Scheme | A standardized framework for classifying recalled information as essential to the storyline (central) or enriching contextual information (peripheral) [15]. | Essential for qualitative analysis of verbal recalls in Protocol 2, particularly for comparing age groups. |
| Semantic Network Analysis Tools | Software pipelines (e.g., NLP libraries in Python/R) to transform narratives into graphs and compute metrics like semantic similarity and centrality [15]. | Used in Protocol 2 to quantify the structural properties of the encoded event and predict recall. |
| Online Experiment Builder | A platform for creating and deploying behavioral experiments (e.g., Gorilla SC, PsychoPy) [15]. | Facilitates precise presentation of stimuli and collection of responses, especially for remote testing. |
Application Notes
This document provides a detailed framework for investigating Temporal Binding Windows (TBWs) in episodic memory. It outlines the core theoretical principles, standardized experimental protocols, and essential research tools for studying how the brain binds discontinuous perceptual elements into unified memory representations across development and experience.
Theoretical Foundation: The Spatiotemporal Scaffold of Memory
Episodic memory relies on binding disparate event details ("what") into a coherent spatiotemporal context ("where" and "when") [18]. Recent theoretical and neural evidence positions the hippocampus and related medial temporal lobe structures as providing a spatiotemporal framework for this process [18] [19].
- The Role of Event Boundaries: Perceptual or contextual shifts, known as event boundaries, play a crucial role in segmenting continuous experience and structuring memory [19]. The Position Coding Model posits that event boundaries affect temporal order memory by resetting the local position coding of events, creating distinct episodic units [20].
- Neural Evidence for Boundary Representations: Intracranial EEG studies reveal that the medial parietal lobe (MPL), including the precuneus, shows unique sensitivity to task boundaries. At boundaries, gamma band activity "resets," which appears to selectively enhance memory for items that appear early in a new sequence (a primacy effect) [19]. This suggests boundaries help define the contextual framework for incoming information.
Developmental Trajectory of Temporal Binding
The ability to bind temporal context undergoes a prolonged development, maturing later than memory for isolated items ("what") [21].
- Emergence of Spatiotemporal Binding: Nonverbal object-placement tasks with children aged 2-8 show that the binding of "where" and "when" (space-time binding) emerges as early as age 3-4 and precedes the development of full episodic memory (what-where-when) [18]. This supports the hypothesis that space-time binding forms the initial scaffold onto which other event details are integrated.
- Refinement with Age: Behavioral and event-related potential (ERP) studies confirm continued age-related improvements in the ability to accurately place events in temporal context throughout middle and late childhood (ages 7-12) [21].
Experimental Protocols
The following protocols are designed to probe the cognitive and neural mechanisms of temporal binding within and across event boundaries.
Protocol 1: Assessing Temporal Order Memory Across Boundaries
This protocol is adapted from paradigms used to investigate the effects of event boundaries on sequence memory [20] [22].
Table 1: Key Experimental Parameters for Temporal Order Memory Task
| Parameter | Specification | Function |
|---|---|---|
| Stimuli | 25 trial-unique images of faces and nameable objects | Provides distinct "what" information and allows for category-based context shifts. |
| Event Boundaries | Semi-predictable shifts in stimulus category and task (e.g., every 2 or 7 items). | Manipulates contextual stability to create within-event and across-event binding conditions. |
| Task During Encoding | Category-specific judgment (e.g., likability for faces, commonality for objects) on a 1-4 scale. | Ensures deep, goal-oriented encoding. |
| Primary Memory Test | Serial recall in order after a distractor period. | Directly measures temporal order memory. |
Detailed Workflow:
- Participant Setup: Secure informed consent. For neuroimaging (fMRI), position the participant in the scanner.
- Encoding Phase:
- Present stimuli sequentially for 2 seconds each.
- Following each stimulus, present a 2-second response period for the category-specific judgment.
- Introduce event boundaries by changing the stimulus category and corresponding task according to a pre-determined schedule.
- Distractor Task: Administer a 45-second arrow discrimination task to prevent rehearsal.
- Recall Phase: Instruct the participant to verbally recall the list of images in the exact order they were presented, skipping items they cannot remember. Record all responses.
Protocol 2: Nonverbal Spatiotemporal Binding in Development
This protocol uses a first-person, 3D real-world task suitable for young children and atypical populations [18].
Table 2: Key Experimental Parameters for Object-Placement Task
| Parameter | Specification | Function |
|---|---|---|
| Stimuli | 3 distinct, small toys and 3 identical opaque containers. | Provides simple, non-verbal "what," "where," and "when" information. |
| Event Structure | Two distinct hiding events, separated by a brief delay. | Creates a temporal sequence for binding. |
| Testing Conditions | Space-Time, Object-Time, and Full Episodic Memory tests. | Dissociates specific memory binding components. |
| Control Condition | "Gated" path during retrieval. | Controls for the role of pure motor memory. |
Detailed Workflow:
- Setup: Place three containers in a triangular arrangement in a room.
- Encoding Phase (Hiding Events):
- Event 1: Show the child a toy (Toy A). Have them walk with the experimenter to a specific container and hide the toy inside. Repeat for a second toy (Toy B) in a different container.
- Delay: Engage the child in an unrelated activity for a short period (e.g., 1-2 minutes).
- Event 2: Show the child a third, distinct toy (Toy C) and have them hide it in the final container.
- Retrieval Phase (Testing):
- Space-Time Test: Ask the child, "Which hiding place did we go to first?" The child must point to or choose the correct container. This assesses the binding of spatial location to temporal order.
- Object-Time Test: Show the child the three toys and ask, "Which toy did we hide first?" This assesses the binding of object identity to temporal order.
- Full Episodic Memory Test: Ask the child, "Which toy is in which box, and which one did we hide first?" This assesses integrated what-where-when memory.
- Gated Condition (Control): For a subset of trials, introduce a physical barrier (e.g., a plastic gate) between encoding and retrieval, forcing the child to take a different path. This ensures the task assesses memory, not path retracing.
The Scientist's Toolkit
Table 3: Essential Research Reagents and Materials
| Item/Category | Function in TBW Research | Specific Examples / Notes |
|---|---|---|
| Stimulus Sets | To provide the "what" element of episodic memory and enable the creation of event boundaries. | Celebrity face and nameable object image banks [22]. Sets of distinct, small toys [18]. |
| Behavioral Task Software | For precise presentation of stimuli, recording of responses, and timing control. | E-Prime, PsychoPy, Presentation. Must support synchronization with neuroimaging equipment. |
| Neuroimaging Hardware | To capture the neural correlates of temporal binding and boundary effects. | Functional MRI (fMRI) scanners [22], intracranial EEG (iEEG) systems [19], and ERP systems [21]. |
| Data Analysis Tools | To quantify behavioral performance and neural activity patterns. | Custom scripts for serial recall analysis (Python, R). Connectivity analysis toolboxes (e.g., CONN for fMRI). Spectral analysis tools for iEEG/ERP data. |
Experimental & Theoretical Model Diagrams
Diagram 1: Experimental Workflow for Assessing Temporal Memory
Diagram 2: Theoretical Framework of Boundary Effects on Memory
Temporal binding refers to the cognitive process by which distinct features of an event, such as sensory inputs and their contextual details, are integrated across time to form a coherent episodic memory. This binding mechanism is fundamental for constructing autobiographical experiences that can be accurately recalled. Emerging theoretical frameworks posit that the brain accomplishes this complex task through processes analogous to Bayesian inference and predictive processing, continuously generating and updating internal models of the environment based on temporal regularities and prediction errors. Within episodic memory research, temporal binding tasks have become crucial experimental paradigms for probing how the hippocampus and associated neural networks encode the "when" component of events, alongside the "what" and "where" information. This document outlines the core theoretical principles of these frameworks and provides detailed application notes and protocols for implementing temporal binding research, specifically designed for scientists investigating memory processes and for drug development professionals targeting cognitive disorders.
Core Theoretical Frameworks
Bayesian Inference Models
Bayesian inference models propose that the brain optimally combines prior knowledge (or "priors") with current sensory evidence (the "likelihood") to form a posterior belief about the state of the world, including the temporal structure of events. In the context of temporal binding for episodic memory, this translates to the integration of pre-existing temporal schemas with incoming sequential information to form a unified memory representation.
- Key Principle: The hippocampus is hypothesized to act as a Bayesian inference engine, computing posterior probabilities for temporal associations. This is mechanistically supported by synaptic plasticity rules that incorporate co-activation probabilities, effectively implementing a form of Bayesian-Hebbian learning [23]. In spiking neural network models, this process enables the dynamic context-item coupling essential for forming episodic memories [23].
- Role of Prior Knowledge: The influence of pre-existing semantic connections on the formation of new, creative associations demonstrates the Bayesian integration of old and new information. Neuroimaging studies show that hippocampal activation patterns during successful encoding vary depending on whether the new associations are remotely or closely related to pre-existing knowledge, reflecting the differential weighting of priors [24].
Predictive Processing Models
Predictive processing models extend the Bayesian framework by positing that the brain is a hierarchical prediction machine. It constantly generates top-down predictions about incoming sensory inputs and uses bottom-up prediction errors to update its internal models.
- Key Principle: Temporal binding is achieved through the minimization of temporal prediction errors. When a sequence of events unfolds as predicted, the elements are bound into a coherent episode with minimal computational cost. A violation of temporal expectations (a prediction error) necessitates a more resource-intensive update of the model, which can either strengthen or disrupt binding.
- Connection to Creativity and Insight: The "insight memory advantage" – where insight solutions are better remembered – has been hypothesized to arise from insight-induced prediction errors detected by the hippocampus [24]. This suggests that temporal binding during novel associative formation (a key creative process) is driven by similar predictive processing mechanisms. Enhanced hippocampal activation and functional connectivity with neocortical regions are observed during the successful encoding of creative associations, reflecting the network-wide updating of predictions [24].
The following diagram illustrates the core architecture of this predictive processing loop in the context of temporal binding.
Diagram 1: Predictive Processing of Temporal Events. The hippocampus generates top-down temporal predictions based on prior beliefs. These are compared to bottom-up sensory input, generating a prediction error signal that drives model updating via synaptic plasticity, resulting in a bound episodic memory.
The following tables synthesize key quantitative findings and parameters from research underlying these theoretical frameworks.
Table 1: Key Experimental Findings in Temporal Memory Processing
| Experimental Paradigm | Key Finding | Quantitative Result / Neural Correlate | Theoretical Implication |
|---|---|---|---|
| Item-in-Context Task (Rodent) [23] | Short-term recency bias competes with episodic memory recall. Rats successfully overcome recency to select old-in-context odors. | Successful recall with retention intervals up to 45 minutes. Model: Bayesian-Hebbian plasticity with eligibility traces. | Co-localization of short- and long-term plasticity enables resolution of temporal conflicts. |
| Libet Clock Task (Human) [25] | Intentional binding (temporal attraction between action and effect) is not solely rooted in spatial working memory. | Correlation between outcome binding and spatial 2-back task was weak and inconclusive (r ≈ small, ns in Exp 2). | Suggests temporal binding may involve specialized mechanisms distinct from spatial WM. |
| Creative Association Encoding (Human fMRI) [24] | Hippocampal representational patterns differ for successful memory of creative associations based on pre-existing semantic links. | Higher inter-item hippocampal pattern similarity for remembered vs. forgotten creative associations. Univariate activation increased for remote-relatedness associations. | Bayesian integration of new associations with prior knowledge is supported by distinct hippocampal processing modes. |
Table 2: Computational Parameters in a Spiking Neural Network Model of Episodic Memory
| Parameter Category | Specific Parameter | Value / Description | Function in Model |
|---|---|---|---|
| Network Architecture | Number of Contexts | 2 | Represents different environmental arenas [23]. |
| Number of Odor-Items | 16 | Distinct items to be associated with contexts [23]. | |
| Temporal Parameters | Inter-Stimulus Interval | 200 ms | Simulates rapid succession of item presentation [23]. |
| Reward Delay for Learning | 250 ms | Time for eligibility trace to bridge delay for reward-based learning [23]. | |
| Plasticity Mechanisms | Within-Network Plasticity | Stable Hebbian attractors | Stores long-term context and item information [23]. |
| Between-Network Plasticity | Bayesian-Hebbian with eligibility traces | Learns and updates episodic item-context associations [23]. | |
| Short-Term Plasticity | Synaptic augmentation/depression | Accounts for recency effects on a millisecond-to-second scale [23]. |
Detailed Experimental Protocols
Protocol: Item-in-Context Temporal Binding Task (Adapted from Panoz-Brown et al.)
This protocol is designed to investigate the interaction between short-term recency effects and long-term episodic memory in rodent models, ideal for testing cognitive enhancers.
1. Objective: To assess an animal's ability to form and recall episodic-like memories of odor-in-context associations while managing conflicting short-term recency signals.
2. Materials and Setup:
- Two Distinct Arenas: Serving as Context A and Context B (e.g., different shapes, flooring, lighting).
- Odor Set: 16 unique, non-confusable odors.
- Reward System: Liquid or food reward dispensers.
- Behavioral Tracking: Video recording and automated tracking software.
3. Procedure:
- Habituation: Animals are familiarized with both arenas and the reward delivery mechanism.
- Item-Context Encoding Block:
- Present 8 specific odors in Context A.
- Subsequently, present all 16 odors in Context B.
- This creates a scenario where 8 odors are "old" to Context A but "new" to Context B, while the other 8 are "new" to both contexts.
- Memory Assessment Block (Immediate Recall):
- In Context B, present a pair of odors in rapid succession (200 ms inter-stimulus interval): one that is "old-in-context" (previously presented in B) and one that is "new-in-context" (not previously presented in B).
- The animal is rewarded for selecting the "new-in-context" odor.
- Critically, the "new-in-context" odor typically has higher recency (was presented more recently in the sequence), putting episodic memory (recall of the earlier presentation in B) in conflict with the short-term recency bias [23].
4. Data Analysis:
- Calculate the percentage of correct choices (selecting the "new-in-context" odor).
- Compare performance across different retention intervals (e.g., from immediate recall to 45 minutes).
- Computational modeling can be applied using a spiking neural network with dual plasticity mechanisms to quantify the contribution of short-term vs. long-term synaptic changes to the behavioral output [23].
The workflow for this protocol, from setup to data analysis, is outlined below.
Diagram 2: Item-in-Context Task Workflow. Sequential stages of the rodent behavioral paradigm for probing temporal binding.
Protocol: Libet Clock Task for Human Temporal Binding
This protocol measures the subjective compression of time between a voluntary action and its sensory outcome, known as intentional binding, a key marker of temporal binding in human agency.
1. Objective: To quantify the sense of agency by measuring the temporal binding effect between actions and their outcomes, and to dissociate it from spatial working memory processes.
2. Materials and Setup:
- Libet Clock: A visual display of a clock face with a rapidly rotating hand (e.g., 2.5s per rotation).
- Response Device: A keypress button for the participant.
- Auditory Stimulus: A tone presented as the outcome of the action.
- 2-Back Working Memory Tasks: Separate spatial and visual 2-back tasks for correlational or dual-task designs [25].
3. Procedure:
- Baseline Judgments:
- Tone Judgment: The tone is played at a random time, and the participant reports the clock hand position when they heard it.
- Action Judgment: The participant performs a keypress at will and reports the clock hand position when they felt the intention to act.
- Binding Trial:
- The participant performs a keypress at will, which triggers the tone after a short, fixed delay (e.g., 250 ms).
- The participant then reports the clock hand position either at the time of their action (for action binding) or at the time of the tone (for outcome binding).
- Control Conditions: Trials with no action, or tones triggered by the experimenter, are included.
- Dual-Task Design (Optional): The Libet clock task can be performed during the retention intervals of a spatial 2-back task to load spatial working memory and test its influence on binding [25].
4. Data Analysis:
- Calculating Binding: The binding effect is the difference in time judgment between the baseline trials and the binding trials. A shift in judgment towards the paired event (e.g., action judged closer to tone) indicates binding.
- Statistical Analysis: Use t-tests or ANOVAs to compare binding effects across conditions (e.g., alone vs. under spatial working memory load). Correlations between binding magnitude and 2-back task performance can also be examined [25].
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents and Materials for Temporal Binding Research
| Item Name | Specifications / Example | Primary Function in Research |
|---|---|---|
| Spiking Neural Network Simulation Environment | e.g., NEST, Brian | To implement and test computational models (e.g., Bayesian-Hebbian models) of temporal binding and synaptic plasticity [23]. |
| Bayesian Modeling Software | e.g., PyMC3, Stan | To fit Bayesian inference models to behavioral data and estimate parameters like prior strengths and prediction errors. |
| Electroencephalography (EEG) System | High-density EEG with real-time processing capability | To measure brain oscillations and implement real-time state-dependent stimulation (e.g., using Bayesian Temporal Prediction) [26]. |
| Functional MRI (fMRI) Scanner | 3T or higher with multiband sequences | To assess hippocampal and neocortical activation and functional connectivity during temporal binding and associative memory tasks [24]. |
| Behavioral Arena Setup (Rodent) | Two or more distinct contexts with odor delivery systems | To conduct item-in-context association tasks that dissociate recency from episodic memory [23]. |
| Libet Clock Software | Custom scripts (e.g., PsychoPy) presenting a rotating clock | To measure intentional binding in human participants as a proxy for temporal binding and sense of agency [25]. |
| Eligibility Trace Component (Computational) | A variable implementing a temporally extended synaptic tag | To model reward-based learning with delays in spiking neural networks, crucial for bridging action-outcome intervals [23]. |
| Spatial Filter (for EEG) | Laplacian montage or source localization algorithm | To accentuate localized brain signals from specific regions of interest (e.g., motor cortex) for real-time phase prediction [26]. |
Assessing Temporal Binding: Experimental Paradigms and Psychopharmacological Applications
The investigation of episodic memory—the ability to recollect the temporal and spatial context of past experiences—relies heavily on behavioral tasks that probe the fundamental mechanisms of temporal processing. Among these, Simultaneity Judgment (SJ) and Temporal Order Judgment (TOJ) tasks serve as core paradigms for quantifying audiovisual temporal perception. These tasks provide a critical window into the temporal binding processes that underlie the formation of coherent episodic memories, where the hippocampus binds item- and context-related information into a unified representation [27]. Research consistently demonstrates that the ability to bind "where" and "when" information emerges early in development and provides a foundational scaffold for full episodic memory, highlighting the centrality of temporal processing in memory formation [18]. The precision of temporal perception, measured through the Temporal Binding Window (TBW), is not only a key individual difference variable but also a malleable target for interventions aimed at improving cognitive and perceptual function [28].
Task Fundamentals and Theoretical Background
Definitions and Core Concepts
- Simultaneity Judgment (SJ): In an SJ task, participants are presented with pairs of crossmodal stimuli (e.g., auditory and visual) at varying stimulus onset asynchronies (SOAs) and are simply asked to report whether the stimuli occurred at the same time or not. The output is a psychometric function describing the proportion of "simultaneous" responses at each SOA [29] [30].
- Temporal Order Judgment (TOJ): In a TOJ task, participants are presented with the same type of stimulus pairs but are asked to indicate which modality appeared first. The output is a psychometric function describing the proportion of "vision first" or "sound first" responses at each SOA [29] [30].
- Temporal Binding Window (TBW): The TBW represents the range of SOAs within which individuals are highly likely to integrate multisensory stimuli into a single perceptual event. A narrower TBW indicates superior audiovisual temporal acuity [28]. The TBW is typically defined as the range of SOAs in which participants perceive simultaneity in a certain percentage (e.g., 75%) of trials or perform at a defined accuracy level on a TOJ task [31].
- Point of Subjective Simultaneity (PSS): The PSS is the specific SOA at which participants are equally likely to judge two stimuli as simultaneous or not, or equally likely to judge either modality as coming first. It represents the perceived timing alignment between sensory modalities and can differ from physical simultaneity (0 ms SOA) due to perceptual biases [31].
Key Quantitative Parameters from the Literature
Table 1: Typical performance metrics and effects of training on SJ and TOJ tasks.
| Parameter | Baseline Performance (Pre-Training) | Post-Training Performance | Key References |
|---|---|---|---|
| Mean TBW Size | ~400-409 ms | Reduces to ~345 ms after 3 days of SJ training | [28] |
| TBW Reduction with Training | N/A | Significant narrowing (58 ms mean decrease; p=0.030) | [28] |
| Training Effect on Reaction Time | N/A | Decreased by 112 ms (p=0.019) after 3 days | [28] |
| Spatial Generalization | N/A | Training in one hemifield completely generalizes to the untrained hemifield | [31] |
| Relationship to Speech-in-Noise | N/A | Larger TBW decreases correlate with improved auditory word recognition (R²=0.291, p=0.038) | [28] |
Relationship to Episodic Memory Framework
The CB (Contextual Binding) theory of episodic memory posits that the hippocampus provides a spatiotemporal framework necessary for representing events, binding together item- and context-related information [27] [18]. According to this view, the ability to accurately perceive and bind temporal sequences through SJ and TOJ tasks reflects a fundamental cognitive process that enables the formation of coherent episodic memories. The developmental trajectory shows that the binding of "where" and "when" emerges around age 3-4 years, preceding and potentially enabling full episodic memory formation, which continues to develop through later childhood [18]. This positions SJ and TOJ tasks as crucial behavioral tools for investigating the temporal component of the spatiotemporal context hypothesized to form the scaffold of episodic memory.
Experimental Protocols
Standardized Simultaneity Judgment (SJ) Protocol
Objective: To determine the range of stimulus onset asynchronies (SOAs) over which an individual perceives audiovisual stimuli as simultaneous and to calculate their Temporal Binding Window (TBW).
Materials and Setup:
- Stimuli: Common stimuli include simple flashes and beeps, or ecological stimuli such as audiovisual speech (e.g., a person saying a syllable) or performing musicians [28] [30].
- Apparatus: The experiment can be conducted in a lab setting or online. Stimuli are presented via a computer screen and headphones. Software such as Psychtoolbox for MATLAB, PsychoPy, or similar experiment builders is required for precise timing control [29].
- SOA Range: A range of SOAs is typically used, from large negative (e.g., sound first) to large positive (vision first) values, often spanning several hundred milliseconds. Common ranges include ±500ms.
Procedure:
- Instruction Phase: Participants are instructed: "You will see a visual stimulus and hear a sound. Your task is to judge whether they happened at exactly the same time. Press 'S' for simultaneous and 'N' for non-simultaneous."
- Trial Structure: Each trial follows this sequence:
- SOA Sampling: Multiple trials are presented for each SOA level. The order of SOAs should be randomized or interleaved to prevent predictability.
- Session Structure: A testing session typically consists of 2-4 blocks of 100-200 trials each. In training studies, sessions are repeated over multiple days (e.g., 3 consecutive days) [28].
Data Analysis:
- Psychometric Function Fitting: For each participant, the proportion of "simultaneous" responses is plotted as a function of SOA. A Gaussian function or a cumulative Gaussian is often fitted to this data.
- TBW Calculation: The TBW is typically defined as the width of the fitted psychometric function at a certain threshold, for instance, the range of SOAs for which simultaneity is reported in 75% of the trials or more [31].
- PSS Calculation: The Point of Subjective Simultaneity is derived as the mean (peak) of the fitted Gaussian function.
Standardized Temporal Order Judgment (TOJ) Protocol
Objective: To determine the SOA at which an individual can reliably discriminate the order of two crossmodal stimuli and to calculate their Just Noticeable Difference (JND).
Materials and Setup: Largely identical to the SJ protocol.
Procedure:
- Instruction Phase: Participants are instructed: "You will see a visual stimulus and hear a sound. Your task is to judge which one came first. Press 'V' for vision first and 'A' for audio first."
- Trial Structure: The trial structure is similar to the SJ task, but without simultaneity feedback in the basic version. Feedback on correctness can be added for training purposes.
- SOA Sampling: The range of SOAs used may be narrower than in SJ tasks, focusing on the region around physical simultaneity where the task is most difficult.
Data Analysis:
- Psychometric Function Fitting: For each participant, the proportion of "vision first" responses is plotted as a function of SOA. A sigmoidal function (e.g., cumulative Gaussian) is fitted to the data.
- PSS Calculation: The PSS is defined as the SOA corresponding to the 50% point on the fitted function, indicating the point of subjective equality.
- JND Calculation: The JND (a measure of temporal acuity) is typically calculated as half the difference between the SOAs corresponding to the 25% and 75% points on the psychometric function. A smaller JND indicates better temporal order perception.
Experimental Workflow for a Typical Study
The following diagram illustrates the standard workflow for implementing and analyzing SJ and TOJ tasks in a research setting.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential materials, software, and tools for implementing SJ and TOJ protocols.
| Category | Item / Solution | Function / Application | Example / Specification |
|---|---|---|---|
| Stimulus Presentation | Presentation Software | Precise control of stimulus timing and sequence | PsychoPy, Psychtoolbox (MATLAB), E-Prime, OpenSesame |
| Audiovisual Stimuli | The core sensory inputs for temporal judgment | Simple (Flash-Beep) or Ecological (Audiovisual Speech) [28] [30] | |
| Response Collection | Input Device | Recording participant judgments with minimal latency | Keyboard, Response Box, Touchscreen |
| Stimulus Control | Audiometric Equipment | Calibrated presentation of auditory stimuli | Headphones, Sound-Attentuating Booth, Audio Interface |
| Visual Display | Calibrated presentation of visual stimuli | CRT Monitor (for minimal lag) or high-refresh-rate LCD | |
| Data Analysis | Statistical Software | Psychometric function fitting and statistical testing | R, Python (with SciPy/Statsmodels), MATLAB, SPSS |
| Neural Correlates | Neuroimaging Techniques | Linking behavioral performance to brain activity | fNIRS (compatible with hearing devices), fMRI, EEG [28] |
| Computational Modeling | Multisensory Correlation Detector (MCD) Model | A stimulus-computable model to simulate and understand perception based on low-level audiovisual input [30] | Population model that processes raw pixels and audio samples |
Key Methodological Considerations
Paradigm Selection: SJ vs. TOJ
The choice between SJ and TOJ paradigms is critical and depends on the research question. A large-scale synthesis of 185 studies reveals that these two paradigms differ not only in task demands but also in their reported outcomes, fueling ongoing debate about their comparability and validity [29] [32]. The SJ task is often considered a more direct measure of the perceived temporal structure of an event, while the TOJ task requires a higher-level categorical decision about order. It is important to note that methodological heterogeneity, particularly in the sampling of temporal lags and testing environment (lab vs. online), has been a major source of contradictory findings in the literature. Adopting a harmonized analytic framework is essential for drawing general conclusions [29].
Training and Plasticity Protocols
Multisensory temporal acuity, as indexed by the TBW, is malleable and can be enhanced through targeted training. A proven protocol involves:
- Structure: 3 consecutive days of SJ training with feedback [28].
- Trials: Multiple blocks of SJ trials per day.
- Feedback: Immediate correctness feedback after each trial ("Correct"/"Incorrect").
- Outcome: This regimen leads to a significant narrowing of the TBW, which is associated with functional benefits such as improved auditory word recognition in noise for individuals who show the largest TBW decreases [28]. Furthermore, this learning completely generalizes to untrained spatial locations, suggesting plasticity at higher, location-invariant processing stages [31]. This has promising implications for clinical interventions aimed at improving temporal processing in populations with enlarged TBWs (e.g., autism, schizophrenia).
Integration with Episodic Memory Assessment
To directly link SJ/TOJ performance to episodic memory within the Contextual Binding theory framework, researchers can:
- Correlational Designs: Administer SJ/TOJ tasks alongside nonverbal episodic memory tasks that dissociate the "what," "where," and "when" components of memory [18].
- Longitudinal Studies: Track the development of TBW narrowing and the emergence of space-time binding in early childhood (ages 2-4), which is hypothesized to scaffold later episodic memory abilities [18].
- Neuroimaging: Use compatible neuroimaging techniques like fNIRS to simultaneously measure hippocampal activity or plasticity in multisensory cortical networks (e.g., left middle temporal gyrus, angular gyrus) during temporal judgment tasks, linking brain function to behavior [27] [28].
{article content start}
Electrophysiological and Eye-Tracking Measures of Memory-Guided Saccades
Memory-guided saccades (MGS) offer a unique window into the cognitive and neural processes underlying episodic memory. This protocol details the application of combined eye-tracking and electrophysiological measures, particularly electroencephalography (EEG), to investigate temporal binding in episodic memory research. We provide a comprehensive methodology for quantifying oculomotor behavior and neural oscillations during MGS tasks, summarizing key quantitative findings and offering a detailed experimental workflow. This resource is designed to equip researchers and drug development professionals with robust tools for assessing hippocampal-dependent memory function and its pathologies.
Episodic memory, the ability to recall personally experienced events anchored in time and place, relies critically on the hippocampus and related medial temporal lobe structures for binding disparate event elements into cohesive representations [7] [33]. Temporal binding—the integration of "what," "where," and "when" information—is a core process in episodic memory formation and retrieval [34].
Memory-guided saccades, where eye movements are made to the remembered location of a visual target after its disappearance, provide a sensitive behavioral readout of this binding process. The accuracy and dynamics of MGS are linked to spatial working memory and the integrity of frontoparietal networks [35]. Furthermore, when combined with electrophysiological recordings such as EEG, MGS tasks can reveal the neural dynamics of memory retrieval. Research shows that successful relational memory retrieval is characterized by a specific neural signature: a transient increase in theta power (4-8 Hz) and a sustained decrease in alpha power (8-13 Hz) [36]. These oscillations are thought to reflect, respectively, the recollection process and the sustained reactivation of goal-relevant associations.
This application note integrates these approaches, providing standardized protocols for using MGS and EEG to probe the temporal binding mechanisms of episodic memory.
The following tables consolidate key quantitative findings from recent research, providing a reference for expected outcomes and effect sizes.
Table 1: Eye-Tracking Metrics in Memory-Guided Saccades and Their Cognitive Correlates
| Metric | Definition | Correlation / Finding | Associated Test/Condition | Source |
|---|---|---|---|---|
| MGS Accuracy | Precision of saccade landing position relative to remembered target location. | Negatively correlated with age (ρ = -0.274). | Memory-Guided Saccade Task (MGST) | [35] |
| Late Errors | Errors in the final saccade position. | Positively correlated with age (ρ = 0.327). | Memory-Guided Saccade Task (MGST) | [35] |
| Corrective Saccade Latency | Speed of initiating a corrective saccade to the actual target. | Positively correlated with MoCA score (ρ = 0.259). | Memory-Guided Saccade Task (MGST) | [35] |
| Early Errors | Erroneous saccades initiated during the memory delay period. | Positively correlated with HADS-A (ρ = 0.325) and HADS-D (ρ = 0.311). | Memory-Guided Saccade Task (MGST) | [35] |
| Gaze Time on Target | Percentage of total gaze time spent on the target vs. a lure. | Most important feature for predicting retrieval accuracy at the single-trial level. | Visual Object Association Task | [37] |
| Trial Entropy | A measure of the disorganization or chaos in scanning patterns. | Key predictive factor for diagnosing temporal lobe epilepsy (TLE) vs. healthy controls. | Visual Object Association Task | [37] |
Table 2: Electrophysiological Correlates of Successful Relational Memory Retrieval
| Oscillation Band | Functional Role | Observed Change During Successful Retrieval | Temporal Characteristics | Source |
|---|---|---|---|---|
| Theta (4-8 Hz) | Recollection process, pattern completion, coordination of sequential information. | Transient power increase. | Emerges shortly after initial cue fixation; predicts success regardless of subsequent saccade target. | [36] |
| Alpha (8-13 Hz) | Reactivation of memory content, cortical engagement. | Sustained power decrease. | Predicts retrieval success specifically when fixations continue on the target; reflects goal-relevant association reactivation. | [36] |
Experimental Protocols
Memory-Guided Saccade Task (MGST) for Cognitive Assessment
This protocol is adapted from a clinical study on stroke patients, providing a method for assessing spatial working memory and executive function [35].
A. Equipment and Setup
- Eye Tracker: A high-quality eye tracker (e.g., Tobii TX300) with a sampling rate of at least 250 Hz.
- Stimulus Presentation: Software such as Tobii Studio or equivalent (e.g., PsychoPy, E-Prime).
- Display: A monitor with a resolution of 1920 × 1080 and a refresh rate of 60 Hz or higher.
- Environment: A quiet, dark room to minimize distractions.
- Participant Setup: Participants are seated 65 cm from the screen with their head stabilized by a chin and forehead rest.
B. Stimulus and Task Design
- Stimulus: A central fixation dot (visual angle of 0.4°) and a peripheral target dot of the same size are presented on a black background.
- Trial Structure:
- Central Fixation: Participant fixates on the central dot for a variable interval (e.g., 1000-1500 ms).
- Target Flash: A peripheral target is briefly flashed (e.g., 100-250 ms) while the participant must maintain central fixation.
- Memory Delay: The central fixation remains visible for a delay period (e.g., 1500-3000 ms). The participant must continue fixating centrally and suppress reflexive saccades.
- Go Signal: The central fixation dot disappears. This is the cue for the participant to make a saccade as accurately as possible to the remembered location of the peripheral target.
- Confirmation Interval: After the MGS is completed, the actual target reappears at its original location, allowing the participant to make a corrective saccade. This confirms the target location and provides feedback.
C. Data Processing and Analysis
- Fixation Filtering: Use a built-in or custom algorithm (e.g., I-VT filter) to classify fixations and saccades. Common parameters include a velocity threshold of 30°/s and a minimum fixation duration of 60 ms [35].
- Trial Validation: Exclude trials with blinks, anticipatory saccades during the delay, or poor calibration.
- Key Metrics:
- MGS Accuracy: Angular or pixel error between the landing position of the first saccade after the "go signal" and the actual target location.
- Early/Late Errors: Rates of saccades made during the delay period (early) or errors in final position (late).
- Corrective Saccade Latency: Time from the "go signal" to the initiation of the first corrective saccade after target reappearance.
Combined EEG and Eye-Tracking for Relational Memory Retrieval
This protocol details a method for capturing the neural dynamics of memory retrieval, as demonstrated in a study on relational memory [36].
A. Equipment and Setup
- EEG System: A high-density EEG system (e.g., 64-channel) with a high sampling rate (≥ 1000 Hz).
- Eye Tracker: A screen-based or head-mounted eye tracker synchronized with the EEG system.
- Stimuli: Hundreds of unique images of faces, places, and objects to prevent repetition effects.
B. Task Design: Associative Memory Test
- Study Phase: Participants view a series of "event" screens, each containing multiple elements from different categories (e.g., 2 faces, 2 places, 2 objects) presented for 10 seconds. They are instructed to memorize all associations within the event.
- Distractor Task: A brief (e.g., 30-second) arithmetic task (e.g., counting backward) is used to clear working memory before the test phase.
- Test Phase (Two-Alternative Forced-Choice):
- Each trial presents a cue element, a target (from the same event as the cue), and a distractor (from a different event).
- The target and distractor are from the same category.
- The participant must fixate freely and select which of the two alternatives was part of the same event as the cue. The test screen is typically presented for 2 seconds.
C. Data Processing and Analysis
- Preprocessing: Standard EEG preprocessing (filtering, bad channel removal, artifact rejection, independent component analysis to remove eye movement and blink artifacts).
- Time-Frequency Analysis: Compute time-frequency representations of power for theta and alpha bands. Analyses are time-locked to stimulus onset and to fixation events.
- Gaze-Contingent Analysis: Segment EEG data based on the sequence of fixations (e.g., cue fixation, target fixation, distractor fixation). Compare neural activity preceding and during fixations on targets versus distractors.
Visualizations
MGS and EEG Experimental Workflow
Neural & Oculomotor Signatures of Retrieval
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Materials and Tools for MGS and EEG Episodic Memory Research
| Item | Function & Application in Research | Example/Specifications |
|---|---|---|
| High-Density EEG System | Records electrophysiological brain activity (e.g., theta/alpha oscillations) with high temporal resolution during memory retrieval. | 64+ channels, sampling rate ≥ 1000 Hz. |
| Research-Grade Eye Tracker | Precisely measures saccades, fixations, and pupil size to quantify oculomotor behavior during memory tasks. | Tobii TX300, Eyelink 1000; sampling rate ≥ 250 Hz. |
| Stimulus Presentation Software | Controls the precise timing and presentation of visual paradigms (e.g., MGST, associative memory tasks). | MATLAB with Psychtoolbox, PsychoPy, E-Prime, Tobii Studio. |
| Visual Object Stimulus Sets | Standardized sets of images used in associative memory tasks to ensure reproducibility and control for low-level features. | Databases of grayscale photos of faces, places, and objects [36]. |
| Cognitive Assessment Batteries | Standardized neuropsychological tests used to correlate eye-tracking/EEG metrics with overall cognitive function. | Montreal Cognitive Assessment (MoCA), Color Trails Test, Digit Span Test [35]. |
| Memory-Guided Saccade Paradigm | The core behavioral task used to probe spatial working memory and frontoparietal network function. | Protocol involving central fixation, target flash, memory delay, and memory-guided saccade [35]. |
| Associative Memory Task | A paradigm designed to test relational (episodic) binding, often used in combination with EEG. | Two-alternative forced-choice task with multi-element event encoding [36]. |
The integration of electrophysiological and eye-tracking measures provides a powerful, multi-dimensional approach to investigating memory-guided saccades within the context of temporal binding in episodic memory. The protocols and data summarized here offer a framework for objectively quantifying memory function, with significant applications in basic cognitive neuroscience, clinical assessment of memory disorders, and pharmaceutical research targeting cognitive enhancement.
{article content end}
Psychoactive Drug Effects on Emotional Episodic Memory Encoding, Consolidation, and Retrieval
Episodic memory, the ability to recollect the temporal and spatial context of past experiences, is a cornerstone of human cognition [27]. Emotional episodic memories, in particular, are vital for adaptive behavior and are significantly modulated by various psychoactive drugs [38]. Understanding these drug effects is crucial not only for fundamental memory research but also for therapeutic drug development. This review is framed within the context of a broader thesis investigating temporal binding tasks for episodic memory research. The Contextual Binding (CB) theory provides a powerful alternative to traditional Systems Consolidation (SC) theory, positing that the hippocampus provides a spatiotemporal framework for representing events and that forgetting is largely due to contextual interference rather than failed consolidation [27]. This theoretical framework is essential for interpreting how psychoactive drugs might alter the binding of item and context information, thereby distorting or enhancing the temporal structure of memories.
Theoretical Framework: Contextual Binding in Episodic Memory
The CB theory posits that the hippocampus binds item- and context-related information, forming the core of an episodic memory [27]. According to this view, memory remains dependent on the hippocampus across time, and forgetting occurs due to contextual interference as the mental and physical context of an individual gradually changes. This is in direct contrast to the Systems Consolidation theory, which suggests memories are transferred from the hippocampus to the neocortex for long-term storage [27]. For research on psychoactive drugs, the CB framework suggests that drug effects should be examined not only on memory items but also on the contextual scaffold that binds those items, including temporal sequence and spatial setting. Drug-induced alterations could disrupt or enhance this binding process, thereby affecting the coherence and accuracy of the retrieved memory.
Acute Effects of Psychoactive Drugs on Emotional Episodic Memory
A comprehensive review of the literature reveals that psychoactive drugs exert distinct, stage-dependent effects on emotional episodic memory [38]. These effects are summarized quantitatively in Table 1.
Table 1: Acute Effects of Psychoactive Drugs on Emotional Episodic Memory Stages
| Drug Class | Specific Agent | Encoding | Consolidation | Retrieval |
|---|---|---|---|---|
| GABAA Sedatives | Alcohol, Benzodiazepines, Zolpidem | Preferentially impairs emotional memory [38] | Preferentially enhances emotional memory (effect varies with delay) [38] | Distorts memory (potentially greater for emotional) [38] |
| Cannabinoids | Δ9-Tetrahydrocannabinol (THC) | Preferentially impairs emotional memory [38] | Information Not Specified | Distorts memory [38] |
| Stimulants | Dextroamphetamine, Dextromethamphetamine | Preferentially enhances emotional memory [38] | Information Not Specified | Distorts memory [38] |
| Dissociative Anesthetics | Ketamine | Preferentially impairs emotional memory [38] | Information Not Specified | Information Not Specified |
| Empathogens | ± 3,4-Methylenedioxymethylamphetamine (MDMA) | Both impairs and enhances (varies by valence) [38] | Information Not Specified | Distorts memory [38] |
Signaling Pathways and Neurobiological Mechanisms
The effects outlined in Table 1 are mediated by interactions with key neurotransmitter systems. The following diagram illustrates the primary molecular targets and the subsequent impact on memory-related brain processes.
Experimental Protocols for Assessing Drug Effects on Temporal Memory Binding
To investigate the effects of psychoactive drugs on episodic memory within the temporal binding framework, specific, robust experimental paradigms are required. The following protocols are adapted from cognitive neuroscience research to be suitable for psychopharmacological studies.
Protocol: Nonverbal Object-Hiding Task for Spatiotemporal Binding
This protocol is ideal for assessing the core premise of the CB theory—the binding of items to a spatiotemporal context—and is easily adaptable for drug studies [18].
- Objective: To assess the differential effects of a psychoactive drug on the binding of object (what), location (where), and temporal sequence (when) information into a coherent episodic memory.
- Materials:
- A testing room with a defined 3x3 grid on the floor.
- 6 distinct, emotionally neutral or valenced objects.
- A plastic gate (for the "gated condition" control).
- Video recording equipment.
- Procedure:
- Encoding Phase (Pre-Drug Administration): The participant is shown three distinct objects being hidden in three different locations of the grid in a specific sequence. The participant is allowed to move freely in the space during this phase.
- Drug Administration: The participant receives the active drug or placebo in a double-blind, randomized design.
- Consolidation Period: A delay of 1-4 hours is implemented, during which the participant remains in a controlled environment.
- Retrieval Phase (Post-Drug Administration): The participant is asked to retrieve the objects. The test is structured to dissociate memory components:
- Space-Time Binding Test: The participant is asked to point to the locations in the exact order they were hidden.
- Object-Time Binding Test: The participant is shown the objects and asked to recall the order in which they were hidden.
- Full Episodic Memory Test: The participant is asked to place each object in its correct location and in the correct temporal order.
- Data Analysis: Performance is scored separately for each binding component (space-time, object-time, full EM). A significant drug effect on space-time binding would provide strong support for its action on the hippocampal spatiotemporal framework proposed by the CB theory [18].
Protocol: Paired-Associate Learning with Contextual Interference
This protocol directly tests the CB theory's explanation of forgetting by measuring the ability to form complex relational structures under drug influence [39].
- Objective: To evaluate how a psychoactive drug impacts the formation of complex item-context memory structures under conditions of high contextual interference.
- Materials:
- Computerized task presentation software.
- Sets of image pairs (e.g., A-B, C-D, A-C, etc.).
- Distinct list-context cues (e.g., colored houses, cartoon characters).
- Procedure:
- Drug Administration: Participants receive the active drug or placebo.
- Study Phase: Participants sequentially study two lists of item pairs. The list type is manipulated to require different memory structures:
- ABCD Lists: Pairs are unique (A-B, C-D). Requires simple two-way binding (A-B).
- ABAC Lists: Pairs share a common element (A-B, A-C). Requires two two-way bindings (A-B + List 1 context).
- ABABr Lists: Pairs are rearranged (List 1: A-B, C-D; List 2: A-D, C-B). Requires three-way binding (A-List 1-B).
- Test Phase: After a delay, participants undergo a cued-recall test for items from both lists.
- Data Analysis: A Multinomial-Processing Tree (MPT) model is applied to estimate the use of different memory structures (two-way vs. three-way bindings) [39]. A drug that increases susceptibility to interference would show a specific reduction in the use of three-way bindings in the ABABr condition.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Materials for Episodic Memory Psychopharmacology Research
| Item | Function/Justification |
|---|---|
| Pharmacological Agents | Reference standards for psychoactive drugs (e.g., THC, dextroamphetamine, midazolam) and matched placebos for double-blind testing. |
| List Context Cues | Distinct visual cues (e.g., colored houses, cartoon characters) presented during list-learning tasks to provide a stable "context" for binding item information [39]. |
| Emotional Stimuli Sets | Standardized image sets (e.g., IAPS) and word lists with normative ratings for emotional valence and arousal, crucial for testing emotional memory modulation [38]. |
| 3D Object Hiding Apparatus | A physical grid (e.g., 3x3) and a set of unique objects for nonverbal assessment of spatiotemporal memory binding in real-world space [18]. |
| Multinomial Processing Tree (MPT) Models | Statistical modeling tool to deconstruct and quantify the use of specific memory processes (e.g., two-way vs. three-way binding) from behavioral data [39]. |
Integrated Experimental Workflow
Combining the theoretical principles, drug effects, and protocols, the following diagram outlines a comprehensive experimental workflow for a study in this field.
Application Notes: Neuropharmacology of Episodic Memory
Episodic memory, the ability to recall specific autobiographical events, is critically dependent on medial temporal lobe structures, primarily the hippocampus. Research utilizing temporal binding tasks, which investigate how the brain links and separates events in memory, provides a powerful framework for quantifying the effects of various psychoactive substances. The following application notes detail how major drug classes modulate the neural substrates of episodic memory, with direct implications for experimental design and interpretation in cognitive research.
GABAA Receptor Agonists and Positive Modulators
Mechanism and Impact: Drugs enhancing Gamma-aminobutyric acid (GABA)ergic transmission at GABAA receptors can significantly impair hippocampus-dependent memory. Notably, receptors containing the α5 subunit are predominantly extrasynaptic and mediate tonic inhibition. Their activation raises the threshold for Long-Term Potentiation (LTP), a key cellular mechanism for memory formation, thereby impairing learning [40]. Inverse agonism of the α5 subunit, conversely, has been shown to improve spatial learning and object recognition memory in animal models [40].
Evidence from Temporal Lobe Epilepsy: In humans, a PET imaging study using the α5-subunit preferring ligand [11C]Ro15-4513 revealed that individuals with MRI-negative temporal lobe epilepsy and co-morbid memory impairments exhibited a ~24-26% bilateral increase in α5 subunit availability in the hippocampal areas. This was associated with a subunit shift from α1/2/3 to α5, indicating increased tonic inhibition as a potential substrate for memory deficits [40].
Other Sedatives: Beyond targeted α5 modulators, broader GABAA modulators like benzodiazepines induce dose-dependent anterograde amnesia and impair working memory, with effects potentially persisting long after withdrawal [41] [42]. Similarly, the neurosteroid allopregnanolone, a positive GABA-A receptor modulator, has been shown to impair memory and accelerate Alzheimer's pathology in transgenic mouse models when exposure is continuous, though intermittent administration can have the opposite effect [43].
Table 1: Quantitative Effects of GABAA Modulators on Memory and Neural Correlates
| Drug / Condition | Receptor Target | Observed Effect on Memory | Key Neural Change |
|---|---|---|---|
| α5-containing GABAA activation | GABAA (α5 subunit) | Impairs spatial & episodic memory [40] | ↑ Tonic inhibition; ↓ LTP threshold [40] |
| α5-containing GABAA inverse agonist | GABAA (α5 subunit) | Enhances spatial learning & object recognition [40] | ↓ Tonic inhibition [40] |
| Temporal Lobe Epilepsy (Human) | GABAA (α5 subunit shift) | Impaired episodic memory [40] | ↑ α5 availability in hippocampus (~25%); ↓ VF:VS ratio [40] |
| Benzodiazepines | GABAA (broad) | Anterograde amnesia; working memory impairment [41] [42] | Reduced brain activation in prefrontal regions [42] |
| Continuous Allopregnanolone | GABAA (neurosteroid site) | Impairs memory; worsens AD pathology in mice [43] | Enhanced tonic inhibition [43] |
Cannabinoids
Impact on Working Memory: The impact of cannabis on memory is one of the most robust findings in neuropsychopharmacology. A large-scale cross-sectional study (n=1,003) of young adults found that heavy lifetime cannabis use (defined as >1000 uses) was associated with statistically significant reductions in brain activation during a working memory task. Specifically, 63% of heavy users showed lower activation in key cognitive regions, including the dorsolateral prefrontal cortex (dlPFC), dorsomedial prefrontal cortex, and anterior insula. This reduced activation was associated with worse task performance [44] [45].
Acute vs. Chronic Effects: The study differentiated between recent use (detected via urine toxicology) and lifetime use, with 68% of recent users also showing reduced brain activation. This suggests that cannabis exerts acute impairing effects on working memory circuitry, which may persist with chronic, heavy use [44].
Interaction with Nicotine: The cognitive profile of cannabis use is complicated by frequent co-use with nicotine/tobacco products (NTP). A systematic review of 39 studies surprisingly found that people with cannabis-NTP co-use often showed similar working memory performance and brain function to non-users. This suggests that nicotine may offset cannabis-related memory deficits, indicating a potential compensatory rather than additive effect when the substances are used together [46].
Table 2: Quantitative Effects of Cannabinoids on Memory and Neural Correlates
| Drug / Condition | Study Population | Observed Effect on Memory | Key Neural Change |
|---|---|---|---|
| Cannabis (Heavy Lifetime Use) | Young adults (n=1003) | Impaired working memory performance [44] [45] | ↓ Activation in dlPFC, dmPFC, anterior insula (63% of users) [44] [45] |
| Cannabis (Recent Use) | Young adults (n=1003) | Impaired working memory performance [44] | ↓ Activation in working memory network (68% of users) [44] |
| Cannabis & Nicotine Co-use | Systematic Review (39 studies) | Working memory similar to non-users [46] | Brain function similar to non-users (offsetting effects) [46] |
Anticholinergics and Stimulants
Anticholinergics: Blockade of muscarinic M1 receptors is a well-established model for episodic memory impairment. A study on biperiden, an M1 antagonist, specifically examined its effect on the serial position curve in a word learning task. While biperiden had an overall impairing effect on episodic memory, it did not mimic the primacy effect (impaired memory for the first words in a list) deficit seen in Alzheimer's disease. Instead, it specifically impaired memory for the middle items in the list during delayed recall, suggesting that M1 receptors are crucial for encoding or consolidating weakly stored information [47].
Stimulants and Dopamine: Dopaminergic signaling is critically involved in both time perception and memory encoding, making it highly relevant for temporal binding tasks. A preprint study combining fMRI and eye-tracking found that event boundaries (context changes) activated the Ventral Tegmental Area (VTA), a key dopaminergic region. This VTA response predicted greater time dilation in memory for events spanning the boundary. Furthermore, eye blinks (a potential marker of dopaminergic activity) also increased at boundaries and predicted time dilation. This suggests that phasic dopamine release at event boundaries drives the expansion of remembered time, a key temporal distortion that helps separate distinct episodic memories [48].
Table 3: Quantitative Effects of Other Drug Classes on Memory
| Drug / Condition | Receptor Target | Observed Effect on Memory | Key Behavioral/Brain Finding |
|---|---|---|---|
| Biperiden | Muscarinic M1 Receptor | Impairs episodic delayed recall [47] | Specific deficit for middle words in a list; no primacy effect deficit [47] |
| Event Boundaries | Dopaminergic System (VTA) | Time dilation in memory [48] | VTA activation & ↑ blinking predict exaggerated temporal distance estimates [48] |
| Psychostimulants (in Polydrug Users) | Multiple (e.g., Dopamine) | Potential benefit on memory [41] | In PWIS, use was associated with a positive effect on memory scores [41] |
Experimental Protocols
Protocol 1: Assessing GABAA α5 Receptor Density and Memory Correlation Using [11C]Ro15-4513 PET
1. Objective: To quantify the availability of α5-containing GABAA receptors in the human brain in vivo and correlate it with performance on episodic memory tasks.
2. Background: The radioligand [11C]Ro15-4513 has a 10-15 times higher affinity for GABAA receptors containing the α5 subunit compared to those with α1-3 subunits. This protocol uses a bolus injection of the ligand with arterial blood sampling to calculate the volume of distribution, separated via "bandpass" exponential spectral analysis into a fast component (VF, dominated by α1-3) and a slow component (VS, dominated by α5) [40].
3. Materials:
- Radiologand: [11C]Ro15-4513.
- Imaging: PET scanner.
- Blood Sampling: Kit for arterial blood sampling and metabolite correction.
- Memory Assessment: Standardized episodic memory battery (e.g., Adult Memory and Information Processing Battery).
- Analysis Software: Software for voxel-by-voxel parametric mapping of VF and VS.
4. Procedure: 1. Participant Preparation: Screen participants for neurological/psychiatric conditions. Obtain informed consent. 2. Radioligand Injection: Administer [11C]Ro15-4513 as a bolus injection. 3. PET Scanning: Initiate a 90-minute dynamic PET scan at the time of injection. 4. Arterial Blood Sampling: Collect arterial blood throughout the scan period for metabolite correction and input function generation. 5. Memory Testing: Administer the episodic memory battery on the same day as the scan. 6. Data Analysis: - Calculate parametric maps of VF and VS from the PET data. - Perform voxel-by-voxel group comparisons (e.g., patient vs. control). - Extract VS values from hippocampal and temporal lobe ROIs. - Correlate hippocampal VS values with episodic memory scores using linear regression.
Protocol 2: fMRI Investigation of Working Memory Under Cannabis Influence
1. Objective: To measure the impact of recent and lifetime cannabis use on brain activation during a working memory task using functional MRI.
2. Background: This protocol leverages a large-sample design to detect subtle changes in brain function. It defines heavy cannabis use as >1000 lifetime uses and uses urine toxicology to verify recent use. The working memory task reliably engages the dlPFC, dmPFC, and anterior insula [44] [45].
3. Materials:
- Imaging: 3T MRI scanner with fMRI capabilities.
- Cognitive Tasks: A battery of at least 7 cognitive tasks administered during fMRI, including a working memory task (e.g., n-back), reward, emotion, and language tasks.
- Assessment:
- Substance Use: Semi-Structured Assessment for the Genetics of Alcoholism to quantify lifetime use.
- Recent Use: Urine toxicology screen on the day of scanning.
- Analysis Software: fMRI preprocessing and statistical analysis software (e.g., SPM, FSL).
4. Procedure: 1. Participant Screening & Grouping: Recruit a large sample (n>1000) of young adults. Group them into non-users (<10 uses), moderate users (10-999 uses), and heavy users (>1000 uses) based on lifetime use data. 2. Urine Toxicology: Collect a urine sample immediately prior to scanning. 3. fMRI Acquisition: Acquire T1-weighted structural images and T2*-weighted BOLD images during task performance. 4. Task Administration: Present the cognitive task battery in a block or event-related design inside the scanner. For the working memory task, use a contrast that isolates the maintenance/manipulation of information (e.g., 2-back vs. 0-back). 5. Data Analysis: - Preprocess fMRI data (realignment, normalization, smoothing). - Model the BOLD response for the working memory contrast. - Extract mean activation from pre-defined ROIs (dlPFC, dmPFC, anterior insula). - Use linear mixed-effects regression models to assess the association between cannabis use group (and recent use) and brain activation, applying False Discovery Rate (FDR) correction for multiple comparisons.
Protocol 3: Serial Position Effect and M1 Receptor Blockade
1. Objective: To evaluate the specific effects of muscarinic M1 receptor blockade on different components of verbal episodic memory using the serial position effect.
2. Background: Blocking M1 receptors with biperiden impairs episodic memory but does not replicate the primacy deficit of Alzheimer's disease. This protocol uses a word-list learning task to dissect the effect of M1 antagonism on primacy, middle, and recency segments [47].
3. Materials:
- Drug: Biperiden and matched placebo.
- Memory Task: A list of 16 words for auditory presentation.
- Design: Double-blind, randomized, placebo-controlled crossover design.
4. Procedure: 1. Participant Preparation: Healthy adults are recruited and screened. Participants fast before the session. 2. Drug Administration: Administer biperiden or placebo in a double-blind fashion. 3. Encoding: After peak drug plasma concentration is reached, present the list of 16 words auditorily. 4. Recall: - Immediate Recall: Ask participants to recall the words immediately after presentation. - Delayed Recall: After a filled delay (e.g., 20-30 minutes), ask for free recall again. 5. Data Scoring & Analysis: - For each recall test, score the words as: Primacy (first 3 words), Middle (middle 10 words), Recency (last 3 words). - Calculate the proportion of correctly recalled words for each segment. - Use repeated-measures ANOVA to analyze the effects of Drug (biperiden vs. placebo) and Serial Position (primacy, middle, recency) on recall performance. The critical finding is a Drug × Serial Position interaction, with the deficit focused on the middle words in delayed recall.
The Scientist's Toolkit: Research Reagent Solutions
Table 4: Essential Reagents and Materials for Episodic Memory Pharmacological Research
| Research Reagent / Material | Function / Application | Key Characteristics / Notes |
|---|---|---|
| [11C]Ro15-4513 | PET radioligand for imaging α5-containing GABAA receptors. | Selective for α5 subunit; requires arterial input function for kinetic modeling [40]. |
| Biperiden | Muscarinic M1 receptor antagonist for pharmacological challenge. | Used to model specific aspects of episodic memory impairment; does not affect primacy [47]. |
| Standardized Memory Batteries (e.g., AMIPB) | Assessment of verbal and visual episodic memory. | Provides normative data for comparison; essential for correlating brain measures with behavior [40]. |
| fMRI Working Memory Task (e.g., n-back) | Probing neural circuitry of working memory in scanner. | Activates dlPFC, dmPFC, anterior insula; sensitive to cannabinoid effects [44] [45]. |
| Serial Position Word List | Dissecting components of episodic memory encoding and recall. | Allows separation of primacy, middle, and recency effects; sensitive to anticholinergics [47]. |
| Semi-Structured Assessment for Genetics of Alcoholism (SSAGA) | Quantifying lifetime substance use history. | Critical for categorizing participants into user groups (e.g., heavy cannabis use >1000 times) [45]. |
| Event Boundary Paradigm | Investigating temporal distortions in episodic memory. | Uses context shifts to study time dilation; engages dopaminergic VTA pathways [48]. |
Temporal binding windows (TBWs) represent critical time frames during which the brain integrates discrete sensory events into a unified perceptual experience. The malleability of these windows, termed TBW plasticity, presents a promising therapeutic target for conditions characterized by dysfunctional temporal processing, such as schizophrenia, autism spectrum disorder, and age-related cognitive decline. This application note details practical training paradigms and experimental protocols for modifying temporal integration boundaries, enabling researchers to systematically investigate and manipulate these neural mechanisms. The core principle underlying these paradigms is the targeted engagement of spike-timing-dependent plasticity (STDP) mechanisms, which strengthen or weaken synaptic connections based on millisecond-precision timing between pre- and postsynaptic activity [49]. Recent research on Behavioral Time Scale Synaptic Plasticity (BTSP) further illuminates how synaptic changes over seconds-long behavioral timescales can support the one-shot formation of memory traces, directly relevant for episodic memory encoding within temporal contexts [50].
Theoretical Foundations and Key Mechanisms
Neural Plasticity Underlying Temporal Binding
Modifying TBWs requires engaging specific neural plasticity mechanisms that operate across different timescales. Spike-timing-dependent plasticity (STDP) serves as a fundamental cellular mechanism, where repetitive pre- and postsynaptic spike pairs induce long-term changes in synaptic strength when evoked at low frequencies (0.1–5 Hz) [49]. The direction and magnitude of change are critically dependent on the precise time interval between spikes, making it ideal for retraining temporal sensitivity. At the systems level, Behavioral Time Scale Synaptic Plasticity (BTSP) provides a complementary mechanism for one-shot learning of temporal sequences and contexts. Unlike STDP, BTSP does not depend on postsynaptic firing but is gated by stochastic synaptic input from the entorhinal cortex, creating plateau potentials that open a plasticity window lasting several seconds [50]. This extended window enables the integration of temporally dispersed information—a core requirement for forming episodic memories.
Episodic Memory and Temporal Binding
In the context of episodic memory, temporal binding refers to the process of integrating "what," "where," and "when" information into a coherent memory representation [34]. The contextual binding (CB) theory posits that the hippocampus binds item- and context-related information, with forgetting largely resulting from contextual interference rather than failed systems consolidation [27]. This framework directly informs TBW modification approaches, as successful paradigms must facilitate the binding of temporal context into memory traces. The temporal integration boundaries manipulated through TBW plasticity protocols correspond to the timeframes within which these hippocampal bindings are formed and retrieved.
Table 1: Core Neural Plasticity Mechanisms Relevant to TBW Modification
| Mechanism | Timescale | Key Induction Parameters | Functional Role in Temporal Binding |
|---|---|---|---|
| Spike-Timing-Dependent Plasticity (STDP) | Milliseconds | Pre/post spike timing (Δt = ±20ms), 50-100 pairings at 0.1-5 Hz [49] | Fine-temporal coding precision, sensory integration |
| Behavioral Time Scale Synaptic Plasticity (BTSP) | Seconds (1-5s) | Plateau potentials from entorhinal cortex input, single-trial induction [50] | Episodic sequence learning, temporal context binding |
| Contextual Binding Mechanisms | Minutes to hours | Hippocampal-cortical interactions, context drift management [27] | Temporal organization of memories, resistance to interference |
Research Reagent Solutions and Essential Materials
Table 2: Key Research Reagents and Materials for TBW Plasticity Studies
| Item/Category | Specific Examples | Function/Application |
|---|---|---|
| Neuroimaging Platforms | MEG systems (whole-head), fMRI (3T/7T), simultaneous MEG-fMRI setups [51] | Tracking neural dynamics (MEG) and spatial localization (fMRI) during temporal binding tasks |
| Stimulation Equipment | Transcranial Magnetic Stimulation (TMS) with stereotactic navigation, intracortical microstimulation systems | Causal manipulation of specific cortical regions in the episodic memory network [7] |
| Animal Behavior Systems | Automated operant chambers with precision timing, video tracking (e.g., EthoVision), in vivo electrophysiology setups | Implementation of temporal discrimination tasks and simultaneous neural recording in rodent models |
| Computational Tools | MNE-Python for source localization, GPT-2 for stimulus feature extraction, custom transformer encoding models [51] | Building encoding models that combine MEG and fMRI data to estimate latent cortical sources |
| Biochemical Reagents | TMZ for chemotherapy-induced plasticity studies, HDAC inhibitors for epigenetic modulation | Investigating molecular mechanisms of plasticity in disease models (e.g., GBM) [52] |
Quantitative Data Synthesis: TBW Parameters Across Paradigms
Table 3: Quantitative Parameters for TBW Modification Across Species and Paradigms
| Paradigm/Species | Baseline TBW | Optimal Training Duration | Key Stimulus Parameters | Plasticity Outcomes |
|---|---|---|---|---|
| Human Audiovisual Temporal Order Judgment | 50-150ms | 5-10 sessions (45-60min each) | ISI sweep: -200 to +200ms in 25ms steps | 15-30% TBW narrowing measured by ΔJND |
| Rodent Temporal Bisection Task | N/A (behavioral readout) | 10-14 days (20 trials/day) | 2s vs 8s anchor durations, intermediate probes | 20-35% improvement in temporal discrimination threshold |
| Human Speech Comprehension (Naturalistic) | N/A (neural measures) | Single exposure (7+ hours narrative) [51] | Natural speech with temporal features (phonemes, prosody) | Enhanced neural encoding precision in auditory-linguistic cortex |
| BTSP Induction (Rodent CA1) | N/A (synaptic level) | Single-trial (one-shot) [50] | Plateau potential timing (0.5-2s window) | Instantaneous place field formation, content-addressable memory |
Experimental Protocols
Protocol 1: Audiovisual Temporal Discrimination Training
Objective: To narrow the temporal binding window through repeated audiovisual temporal order judgment training.
Materials:
- Presentation system with millisecond precision (e.g., PsychToolbox, E-Prime)
- Audiovisual stimulators with calibrated timing
- EEG/MEG recording system (optional for neural correlates)
- Response collection device
Procedure:
- Baseline Assessment:
- Present audiovisual stimulus pairs with interstimulus intervals (ISIs) from -200ms (sound first) to +200ms (vision first) in randomized order
- For each trial, participant judges whether sound or light appeared first
- Fit psychometric function to determine just noticeable difference (JND) and point of subjective simultaneity (PSS)
- Calculate baseline TBW as ±2 JND around PSS
Training Phase:
- Conduct 5 sessions over 2 weeks (3 days between sessions)
- Each session: 300 trials focused near individual's JND (±1.5 JND from PSS)
- Provide trial-by-trial accuracy feedback with correct response highlighted
- Implement adaptive staircase: after 3 correct, decrease ISI by 5%; after 1 error, increase ISI by 10%
Post-Training Assessment:
- Repeat baseline assessment protocol without feedback
- Compare pre/post JND values and TBW boundaries
Data Analysis:
- Calculate JND and PSS using cumulative Gaussian fit
- Compute TBW narrowing as (preJND - postJND)/pre_JND × 100%
- For neural data: Analyze MEG/EEG components (N1/P1 latencies) and phase-locking values
Figure 1: Audiovisual Temporal Discrimination Training Protocol Workflow
Protocol 2: BTSP-Based Temporal Sequence Learning in Rodents
Objective: To leverage Behavioral Time Scale Synaptic Plasticity for one-shot learning of temporal sequences in rodent models.
Materials:
- Behavioral apparatus with multiple distinct compartments
- In vivo electrophysiology system for CA1 recordings
- Optogenetic stimulation system (if using optogenetic gating)
- Video tracking system with temporal resolution <33ms
Procedure:
- Surgical Preparation (if using neural manipulation):
- Implant recording electrodes in hippocampal CA1 region
- For optogenetic experiments: inject AAV-CaMKIIα-ChR2 in medial entorhinal cortex layer III
- Implant optogenetic fiber above CA1 distal dendrites
BTSP Induction Protocol:
- Habituate animal to behavioral apparatus for 30min/day for 3 days
- On training day: place animal in start compartment, allow exploration
- Trigger sequential activation of compartment cues (A→B→C) with 2s intervals
- During cue sequence, induce plateau potentials via either:
- Natural entorhinal input driven by novel environmental features
- Optogenetic stimulation of entorhinal inputs (5-10ms pulses at 20Hz for 2s)
- Record CA1 place field formation in response to temporal sequence
Memory Testing:
- 24h post-induction: test for place field stability in same environment
- Use partial cue sequences (A→B only) to test pattern completion
- Introduce altered temporal sequences to test temporal specificity
Data Analysis:
- Map place field formation and stability using rate maps
- Calculate spatial information content and temporal specificity index
- Analyze cross-correlations between sequentially activated place cells
Figure 2: BTSP-Based Temporal Sequence Learning Protocol
Protocol 3: Naturalistic Speech Comprehension with Multimodal Imaging
Objective: To measure and manipulate temporal integration boundaries during naturalistic speech processing using MEG-fMRI fusion.
Materials:
- Whole-head MEG system with compatible audio presentation
- 3T/7T fMRI scanner with optimized auditory stimulation
- Naturalistic speech stimuli (narrative stories >7 hours total)
- Transformer-based encoding model infrastructure [51]
Procedure:
- Stimulus Feature Extraction:
- Extract three feature streams from speech stimuli:
- 768-dimensional GPT-2 contextual word embeddings
- 44-dimensional phoneme one-hot vectors
- 40-dimensional mel-spectrograms (0-10 kHz)
- Sample all features at 50Hz temporal resolution
- Extract three feature streams from speech stimuli:
Multimodal Data Collection:
- Collect MEG data during passive listening to narrative stories
- Use identical stimuli in separate fMRI session (blocked design)
- Maintain consistent audio presentation levels across modalities
Encoding Model Construction:
- Build transformer-based architecture with:
- Linear input layer (852 features → 256 dimensions)
- 4 transformer encoder layers with causal sliding window (500 tokens, 10s context)
- Source layer projecting to "fsaverage" source space (8,196 sources)
- Subject-specific source morphing using MNE-Python
- Train model to simultaneously predict MEG and fMRI signals
- Build transformer-based architecture with:
Temporal Integration Analysis:
- Identify neural representations of temporal boundaries in narratives
- Calculate integration windows from cross-regional phase synchronization
- Relate individual differences in TBW to behavioral speech comprehension
Validation with ECoG Data:
- Test generalizability by predicting ECoG responses from estimated sources
- Compare performance against ECoG-trained encoding models
Figure 3: Naturalistic Speech Comprehension with Multimodal Imaging Protocol
Signaling Pathways in TBW Plasticity
Figure 4: Signaling Pathways in Temporal Binding Window Plasticity
Data Analysis and Interpretation Guidelines
Quantitative Metrics for TBW Modification
Table 4: Key Analytical Metrics for Assessing TBW Plasticity Outcomes
| Metric Category | Specific Measures | Calculation Method | Interpretation Guidelines |
|---|---|---|---|
| Psychophysical Performance | Just Noticeable Difference (JND) | Standard deviation of fitted psychometric function | Smaller JND = improved temporal precision |
| Point of Subjective Simultaneity (PSS) | Mean of fitted psychometric function | Shift indicates recalibration of temporal alignment | |
| Neural Timing Precision | Phase-Locking Value (PLV) | Inter-trial coherence of oscillatory phase | Higher PLV = more precise temporal encoding |
| Latency of Evoked Components | Peak times of N1/P1/M1 components | Reduced latency = faster processing | |
| BTSP-Specific Measures | Place Field Specificity | Spatial information content (bits/spike) | Higher values = more precise temporal-spatial binding |
| Temporal Specificity Index | Cross-correlation peak prominence | Higher values = better sequence learning | |
| Network-Level Integration | Functional Connectivity | MEG/EEG source-space coherence | Increased coherence = improved network integration |
Statistical Considerations for TBW Studies
- Sample Size Planning: For within-subjects TBW training studies, target N=20-30 for 80% power to detect moderate effects (d=0.6)
- Multiple Comparison Correction: Use FDR correction for mass univariate analyses (e.g., whole-brain MEG-fMRI)
- Longitudinal Analyses: Employ mixed-effects models to account for within-subject correlations across sessions
- Effect Size Reporting: Always report Cohen's d for TBW narrowing effects alongside p-values
Troubleshooting and Optimization
Common Challenges:
- High variability in training responses: Implement more precise individual baseline assessment and customize training parameters
- Poor retention of TBW narrowing: Include booster sessions at 1-month and 3-month intervals
- Limited generalization across modalities: Train with multiple stimulus types and contexts
- BTSP induction variability: Optimize plateau potential timing and duration based on individual neural responses
Optimization Strategies:
- Systematically vary inter-session intervals (1-5 days) to identify optimal spacing
- Combine peripheral (psychophysical) and central (TMS/tDCS) stimulation approaches
- Use real-time fMRI neurofeedback to reinforce desired neural states during training
- Implement individualized frequency parameters based on baseline oscillatory profiles
Addressing Methodological Challenges and Optimizing Temporal Binding Assessments
Temporal binding, the phenomenon where a voluntary action and its sensory outcome are perceived as closer together in time than they actually are, serves as a crucial implicit measure of the sense of agency [53]. In episodic memory research, this binding effect provides a window into how we form compressed, causal representations of our experiences. However, accurate measurement of temporal binding is compromised by several confounding factors, principally attention, working memory (WM) load, and metacognitive experiences. Attention lapses can distort time perception [54], WM capacity influences the temporal compression of episodic memories [55], and metacognitive failures prevent participants from accurately monitoring their own performance [54]. This application note provides detailed protocols and methodological frameworks to control for these confounds, ensuring the validity and reliability of temporal binding measurements within episodic memory research.
Core Experimental Protocol: The Libet Clock Task for Temporal Binding
The following section details the standard protocol for assessing temporal binding, adapted from established procedures [56] [53].
Apparatus and Stimuli
- Visual Display: Participants view an analogue clock face (diameter: ~16.56° visual angle) marked with numbers at intervals of 5 (0, 5, 10...55). A dot (the clock hand) revolves around the clock. One full rotation should last 2560 ms [56].
- Response Interface: Standard computer keyboard for key presses and a mouse for cursor responses.
- Auditory Stimuli: A tone (1000 Hz, 100 ms duration) can be used as an outcome [56].
- Software: The open-source HTML5 tool Labclock Web is highly recommended for programming and conducting these experiments, including online studies [53].
Detailed Trial Procedure
- Fixation: Participants are instructed to focus on a central fixation cross on the clock face.
- First Rotation (Familiarization): The dot completes one full rotation (2560 ms). Participants observe but do not perform any actions [56].
- Second Rotation (Experimental Phase):
- Participants are instructed to press a key at a time of their own choosing, waiting at least one full revolution before doing so. They are told not to plan or aim for a specific time on the clock [56].
- Depending on the trial type, a key press may trigger a tone after a specific delay (e.g., 250 ms), or a tone may occur randomly.
- Time Estimation: After the second rotation, participants report their time estimation of the target event (either the key press or the tone onset) by clicking the corresponding position on the still-visible clock face with a cursor. Responses should be as precise as possible [56].
- Error Handling: If a participant fails to press the key within the second lap, an error message ("You forgot to press!") accompanied by a sound is displayed for three seconds, prolonging the trial to motivate task engagement [56].
Trial Types and Block Design
To isolate the binding effect from the mere presence of external feedback, a block design with the following trial types is recommended [56] [53]. Each block should contain a sufficient number of trials (e.g., 25 trials plus 3 practice trials [56]).
Table 1: Trial Types in the Libet Clock Task
| Trial Type | Action (Key Press) | Outcome (Tone) | Reported Event | Purpose |
|---|---|---|---|---|
| Agency Action | Yes, triggers tone after 250 ms | Yes | Time of Key Press | Measures shift of action perception toward outcome |
| Agency Outcome | Yes, triggers tone after 250 ms | Yes | Time of Tone | Measures shift of outcome perception toward action |
| Baseline Action | Yes | No | Time of Key Press | Baseline for action perception without outcome |
| Baseline Outcome | No | Yes, random time | Time of Tone | Baseline for outcome perception without action |
Controlling for Confounding Factors
Controlling Attention
Attention lapses, or "off-task" thoughts, are a major source of performance fluctuation and can severely distort time estimations in the Libet task [54].
- Protocol: Probing Subjective Attention State
- Method: On a random subset of trials (e.g., 20-25%), present a thought probe immediately after the time estimation response [54].
- Probe Question: Participants categorize their thought content during the preceding trial into one of four categories [54]:
- On Task: "Totally focused on completing the task."
- Task-Related Interference: "Thinking about task performance."
- Mind-Wandering: "Thinking about something other than the task."
- External Distraction: "Thinking about something in my immediate environment."
- Implementation: The experimenter should provide detailed examples and check for participant understanding before the experiment begins. Responses can be made via keyboard press [54].
- Data Analysis: Trials with reports of mind-wandering or external distraction can be flagged or analyzed separately, as these are associated with significantly worse performance [54].
Controlling Working Memory Load
Working memory capacity is a key determinant of how events are temporally compressed in episodic memory [55]. Manipulating WM load within the temporal binding task allows researchers to directly test its influence.
- Protocol: Integrating a Spatial Working Memory Task
- This protocol is adapted from research on internal attention [57].
- Stimuli: Present an array of colored squares (~2.5° visual angle) on a gray background for 250 ms. Use a set of distinct colors (e.g., red, green, blue, magenta, yellow, cyan, orange, white, black) [54].
- Task: After a blank delay period (e.g., 1000 ms), participants are shown a response grid at the location of each memory item. They must use the mouse to click the color they remember for each location, reporting all items in any order [54].
- Load Manipulation: Implement separate blocks of "easy" (e.g., 2-3 items) and "hard" (e.g., 6-8 items) set-sizes to vary WM load [54].
- Combining with Temporal Binding: The Libet Clock Task can be performed either before the WM encoding period or after the WM retrieval period, depending on whether the research question concerns the effect of WM load on binding, or vice versa.
Controlling Metacognitive Experiences
Participants often lack awareness of their own working memory failures, leading to overconfident judgments on error trials [54]. Assessing metacognition provides insight into this awareness.
- Protocol: Trial-by-Trial Confidence Judgments
- Method: Following the time estimation response on every trial, participants provide a metacognitive judgment [54].
- Implementation: Use a simple, rapid response. For example, present a scale and have participants rate their confidence in the accuracy of their time estimation using a mouse-click. A scale of 1 (Very Uncertain) to 4 (Very Certain) is effective [54].
- Data Analysis: Compute metrics of metacognitive sensitivity (e.g., the correlation between confidence and actual estimation accuracy) for each participant. This allows for the identification of systematic overconfidence on lapse trials.
The following diagram summarizes the core experimental workflow and the points of intervention for controlling confounding factors.
Quantitative Data and Analysis
The table below summarizes key quantitative findings from the literature on the confounding factors discussed, providing a reference for expected effect sizes and outcomes.
Table 2: Key Quantitative Findings on Confounding Factors in Memory and Time Perception
| Factor | Experimental Manipulation | Key Quantitative Result | Implication for Temporal Binding |
|---|---|---|---|
| Attention [54] | Thought probes vs. whole-report WM performance. | Participants identified their own WM failures on only ~28% of failure trials. Off-task thoughts predicted performance. | Lapses in attention likely cause significant noise/error in time estimations, unbeknownst to the participant. |
| Working Memory [57] | Retro-cueing of items in WM vs. LTM. | Attentional orienting benefits were stronger in WM than LTM (Reaction Time: F(1,29)=18.63, p<0.001, η²=0.39; Accuracy: F(1,29)=12.12, p=0.002, η²=0.30). | High WM load may consume resources needed for precise temporal processing, potentially attenuating binding. |
| Working Memory Capacity [55] | Individual differences in WM capacity and mental replay of videos. | WM capacity was negatively associated with temporal compression rates, but only for events with few boundaries. | Higher capacity may lead to more veridical temporal representations, affecting how action-outcome sequences are encoded. |
| Metacognition [54] | Confidence ratings on every trial of a WM task. | Metacognitive judgments reliably predicted performance variation but severely underestimated the extent of failures. | Overconfidence in one's time estimations could be a source of bias, making trial-by-trial confidence crucial. |
The Scientist's Toolkit: Research Reagent Solutions
This table outlines essential materials and tools for implementing the protocols described in this note.
Table 3: Essential Research Tools and Reagents
| Item Name | Function/Description | Example/Specification |
|---|---|---|
| Labclock Web [53] | Open-source HTML5 software for programming and running temporal binding (Libet clock) experiments online or in-lab. | Allows for easy customization of trial parameters, conditions, and stimuli. Essential for standardizing methodology. |
| Whole-Report WM Task [54] [57] | A visual working memory paradigm that provides a graded measure of performance on each trial. | Stimuli: Colored squares (2.5° visual angle). Procedure: 250 ms display, 1000 ms delay, then report all items via mouse click. |
| Attention Probe [54] | A subjective measure to categorize thought content during a trial, identifying attentional lapses. | Four categories: On Task, Task-Related Interference, Mind-Wandering, External Distraction. |
| Confidence Judgment Scale [54] | A simple metacognitive measure to assess a participant's awareness of their performance on each trial. | A 4-point scale from "Very Uncertain" to "Very Certain" provided after the primary task response. |
| Standardized Color Set [54] | A fixed palette of highly distinct colors for visual memory tasks to minimize perceptual confusion. | Example RGB values: Red (255,0,0), Green (0,255,0), Blue (0,0,255), Magenta (255,0,255), Yellow (255,255,0), etc. |
Integrated Experimental Design and Theoretical Framework
To comprehensively control for multiple confounds simultaneously, an integrated design is necessary. The following diagram illustrates the theoretical relationships between the controlled factors and the core phenomenon of temporal binding, framing it within episodic memory research.
Integrated Workflow Recommendation:
- Session Structure: Begin with a WM capacity assessment (e.g., a complex span task) [55].
- Main Task: Conduct the combined Libet Clock Task, embedding:
- Data Analysis: Analyze temporal binding effects (i.e., the shift in time estimation between operant and baseline trials) with WM load as a between-block factor, and include attention probe results and confidence ratings as covariates or factors in the statistical model.
By implementing these controlled protocols, researchers can isolate the specific contributions of attention, working memory, and metacognition to temporal binding, thereby yielding more precise and interpretable results in the study of episodic memory and agency.
Application Notes
The study of episodic memory, the conscious recollection of past experiences, is fundamental to understanding cognitive function and its modulation by psychoactive substances. A critical advancement in this field involves the temporal dissection of memory into its constituent stages: encoding (the initial formation of a memory trace), consolidation (the stabilization of the trace into long-term storage), and retrieval (the access and recall of the stored memory) [58]. For researchers and drug development professionals, isolating the effects of a compound on these distinct stages is paramount for identifying its precise mechanism of action. This is especially relevant within the context of temporal binding tasks, which assess how the brain links the "what," "where," and "when" of an event into a coherent memory [59]. Administering a drug non-specifically before a memory task confounds its potential effects on encoding, consolidation, and retrieval. The protocols herein provide detailed methodologies for decoupling these stages, enabling the isolation of drug effects to a specific mnemonic process and providing a clearer picture of a compound's potential for therapeutic use or abuse [58].
Key Brain Circuits and Molecular Targets
Drugs exert their effects on memory by modulating specific neural circuits and molecular pathways. The limbic–corticostriatal system, comprising structures like the basolateral amygdala (BLA), hippocampus, nucleus accumbens (NAc), and prefrontal cortex, plays a vital role in encoding and reconsolidating drug-related memories [60]. The table below summarizes key molecular targets within these circuits that have been successfully manipulated to disrupt drug memory reconsolidation in rodent models, providing a toolkit for targeted interventions.
Table 1: Key Molecular Targets for Disrupting Drug Memory Reconsolidation
| Brain Area | Drug Used in Model | Molecular Target | Treatment (Example) | Effect on Memory |
|---|---|---|---|---|
| Basolateral Amygdala (BLA) | Cocaine | NMDAR, β-Adrenergic Receptor (β-AR), ERK, Zif268 | D-APV (NMDAR antagonist), Propranolol (β-blocker), U0126 (ERK inhibitor) | Disruption [60] |
| BLA | Morphine | Glucocorticoid Receptors (GRs) | GR agonist | Disruption [60] |
| Nucle Accumbens (NAc) | Cocaine | DNA methyltransferase | RG108 (inhibitor) | Disruption [60] |
| Central Amygdala (CeA) | Alcohol | mTORC1 | Rapamycin (inhibitor) | Disruption [60] |
| Dorsal Hippocampus (DH) | Cocaine | DNA demethylation | Knockdown (Tet3) | Disruption [60] |
| Systemically | Cocaine | mTOR, CB1R | Rapamycin, AM251 (CB1R antagonist) | Disruption [60] |
Experimental Protocols
The following protocols are designed to temporally isolate the stages of memory. They utilize well-established behavioral paradigms such as Conditioned Place Preference (CPP) and Self-Administration (SA) in rodents, which model reward-related learning, and can be adapted for human subjects using episodic memory tasks with emotional and neutral stimuli [60] [58].
Protocol 1: Isolating Drug Effects on Memory Encoding
This protocol isolates the encoding stage by ensuring the drug is active only during memory formation and has been cleared before retrieval, thus eliminating confounding effects on consolidation and retrieval [58].
- Objective: To assess the effects of a drug specifically on the acquisition of new episodic memories.
- Procedure:
- Pre-encoding Phase: Administer the drug or vehicle to subjects.
- Encoding Phase: After the drug has reached peak plasma concentration, expose subjects to the learning material. In rodent studies, this involves a CPP session or an SA training session. In human studies, participants encode stimuli (e.g., images, word pairs) while performing a distractor task to prevent active rehearsal [58].
- Consolidation & Retrieval Delay: Allow a sufficient delay (e.g., ≥24 hours) for the drug to be fully metabolized and eliminated. This ensures the drug is absent during the subsequent consolidation period and retrieval test.
- Retrieval Test: Conduct a drug-free memory test. In rodent CPP, this involves measuring time spent in the drug-paired context. In human studies, this involves free recall or recognition tests for the encoded stimuli [58].
- Key Measurements:
- Rodents: Preference score (time in drug-paired chamber minus time in neutral chamber) in CPP; number of active lever presses in SA.
- Humans: Recall rate/accuracy; recognition sensitivity (d').
- Interpretation: A significant difference in memory performance between the drug and vehicle groups indicates a specific effect on the encoding process. For example, GABAA sedatives (alcohol, benzodiazepines) and THC typically impair encoding, particularly for emotional stimuli [58].
Protocol 2: Isolating Drug Effects on Memory Consolidation
This protocol targets the consolidation stage by administering the drug immediately after the encoding phase, when the memory trace is labile and susceptible to modulation [58] [61].
- Objective: To determine if a drug modulates the stabilization of a memory trace after it has been formed.
- Procedure:
- Encoding Phase: Subjects undergo a drug-free learning session (e.g., CPP training, SA session, or viewing stimuli in humans).
- Post-encoding Phase: Administer the drug or vehicle immediately after the encoding session is complete. The drug must have rapid absorption to be active during the critical window of cellular consolidation.
- Retrieval Delay: Allow for a standard retention interval (e.g., 24 hours) for long-term memory to form.
- Retrieval Test: Conduct a drug-free memory test.
- Key Measurements: Same as Protocol 1.
- Interpretation: A difference in memory performance indicates a drug effect on consolidation. For instance, GABAA sedatives administered post-encoding can retroactively enhance memory consolidation, a phenomenon known as retrograde facilitation, particularly for emotional content [58].
Protocol 3: Isolating Drug Effects on Memory Retrieval
This protocol isolates the retrieval stage by administering the drug after consolidation is complete and just before the memory test [58].
- Objective: To evaluate if a drug affects the access and recall of a stabilized long-term memory.
- Procedure:
- Encoding Phase: Subjects undergo a drug-free learning session.
- Consolidation Period: Allow a standard retention interval (e.g., 24 hours or more) to ensure the memory is fully consolidated and the drug from encoding is absent.
- Pre-retrieval Phase: Administer the drug or vehicle shortly before the retrieval test.
- Retrieval Test: Conduct the memory test while the drug is active.
- Key Measurements: Same as Protocol 1.
- Interpretation: A difference in memory performance indicates a drug effect on the retrieval process. THC, dextroamphetamine, and MDMA have been shown to distort memory retrieval, often making memories more positive or less negative [58].
Signaling Pathways and Experimental Workflows
Neural Circuitry of Memory Reconsolidation
The following diagram illustrates the key brain structures within the limbic–corticostriatal system that interact during drug memory reconsolidation, based on evidence from lesion and pharmacological studies [60].
Diagram 1: Brain circuit of drug memory reconsolidation.
Workflow for Isolating Memory Stages
This workflow outlines the procedural timeline for the three core protocols, highlighting the critical timing of drug administration to isolate effects on encoding, consolidation, and retrieval.
Diagram 2: Workflow for isolating memory stage effects.
The Scientist's Toolkit: Research Reagent Solutions
This table catalogs essential pharmacological tools and their applications for probing the mechanisms of memory stages, particularly in the context of drug memory reconsolidation and emotional episodic memory [60] [58].
Table 2: Key Research Reagents for Memory Mechanism Studies
| Reagent / Agent | Molecular Target | Primary Function | Key Experimental Use |
|---|---|---|---|
| Anisomycin | Protein synthesis | Inhibitor | Disrupts reconsolidation by blocking new protein synthesis required for long-term memory stabilization [60]. |
| Propranolol | β-Adrenergic Receptor (β-AR) | Non-specific β-blocker | Disrupts reconsolidation of cocaine and heroin memories, particularly in the BLA and prefrontal cortex [60]. |
| Rapamycin | mTORC1 | Inhibitor | Disrupts alcohol memory reconsolidation (CeA) and cocaine memory systemically; targets protein synthesis-dependent plasticity [60]. |
| U0126 | ERK (Extracellular signal-regulated kinase) | Inhibitor | Disrupts reconsolidation by blocking the ERK signaling pathway in the BLA [60]. |
| D-APV | NMDAR (N-methyl-D-aspartate receptor) | Antagonist | Disrupts reconsolidation by blocking NMDA receptors, critical for synaptic plasticity, in the BLA [60]. |
| AM251 | CB1R (Cannabinoid type 1 receptor) | Antagonist | Disrupts reconsolidation of cocaine memories when administered systemically [60]. |
| RG108 | DNA methyltransferase | Inhibitor | Disrupts reconsolidation of cocaine memories in the NAc by inhibiting epigenetic mechanisms [60]. |
Temporal binding tasks probe the cognitive mechanisms responsible for integrating disparate features (e.g., colour, shape) into cohesive object representations across time. This process is a fundamental component of episodic memory—the memory for events from a specific time and place [62] [21]. Research indicates that memory for temporal context shows a prolonged developmental trajectory and is subserved by neural structures including the hippocampus and entorhinal cortex [21] [63]. In clinical populations, such as those with Alzheimer's disease, the ability to bind features in visual short-term memory is particularly impaired, making these tasks a promising early marker of pathology [64]. Adapting temporal binding protocols for clinical groups with cognitive and sensory limitations is therefore paramount for accurate assessment and diagnosis. This application note provides a detailed framework for such adaptations, encompassing theoretical foundations, modified experimental protocols, and key methodological considerations.
Theoretical and Neural Basis of Temporal Binding
Temporal binding refers to the process by which features that occur in close temporal proximity are correctly conjoined to form a unified perceptual representation. Failures in this process can lead to temporal binding errors, or illusory conjunctions, where features from successive objects are incorrectly combined [62]. According to the diachronic framework of attention, transient periods of attentional amplification (~150–250 ms), known as "attentional episodes," facilitate the binding of temporally co-occurring features. The speed of attentional engagement is a critical factor in determining binding accuracy [62].
At the neural level, the hippocampus (HPC) and entorhinal cortex (EC) are crucial for processing temporal context. Recent findings demonstrate that intrinsic, spontaneous fluctuations in HPC- and EC-whole-brain functional connectivity patterns systematically drift with the passage of time, serving as a neural signature of temporal context even in the absence of task demands [63]. This intrinsic "time tracker" follows an anterior-to-posterior gradient along the hippocampal longitudinal axis, with the anterior HPC and anterolateral EC showing stronger temporal drifts, suggesting they support coarser, longer-timescale representations [63]. Furthermore, the development of memory for temporal context, a vital aspect of episodic memory, continues to improve across middle and late childhood, as evidenced by event-related potential (ERP) studies [21].
Table 1: Key Neural Correlates of Temporal Context and Binding
| Neural Structure/Measure | Function in Temporal Processing | Relevance to Clinical Adaptation |
|---|---|---|
| Anterior Hippocampus (aHPC) | Tracks temporal information over long timescales; "gist-like" memory [63]. | Vulnerable to age-related decline and neurodegenerative diseases; assessments may need to reduce reliance on long-term recall. |
| Posterior Hippocampus (pHPC) | Supports detailed autobiographical memories; reflects shorter trial-specific intervals [63]. | |
| Entorhinal Cortex (EC) | Represents temporal context; its functional connectivity patterns drift with time [63]. | |
| ERP Old/New Effects | Neural correlates of recognition memory and successful retrieval of temporal source [21]. | Can be used as a neural biomarker in populations where behavioral responses are unreliable (e.g., severe cognitive impairment). |
| Frontal & Occipital Modules | Show differentiated connectivity during VSTM binding tasks, indicating an information processing loop [64]. | Connectivity differences may serve as sensitive biomarkers for diseases like Alzheimer's. |
Core Experimental Protocols and Their Clinical Adaptation
The following section outlines standard protocols for investigating temporal binding and episodic memory, followed by specific adaptations for clinical populations with cognitive and sensory limitations.
The Rapid Serial Visual Presentation (RSVP) Task for Temporal Binding
Original Protocol [62]:
- Objective: To investigate temporal binding errors and the role of attentional engagement.
- Stimuli: A stream of stimuli (e.g., coloured letters or shapes) is presented sequentially at a rapid pace (e.g., 100 ms per item) at a single location.
- Task: Participants are required to report two features (e.g., colour and alphanumeric identity) of a predefined target (e.g., a digit among letters).
- Key Measures: The frequency and pattern of temporal binding errors, particularly the covariance between reported identity and colour. When participants misreport a distractor's identity, they are more likely to misreport that same distractor's colour, indicating a failure of feature binding [62].
Adaptations for Clinical Populations:
- Cognitive Limitations (e.g., ADHD, Dementia):
- Reduce Attentional Load: Use shorter RSVP streams and incorporate more frequent breaks to mitigate fatigue and sustained attention deficits.
- Simplify Response Demands: Instead of requiring two separate verbal or typed responses, use a recognition-based response screen. Present all possible target and distractor colour-identity combinations, along with a "foil" object not in the stream, allowing participants to select the item they believe was the target [62]. This reduces the load on working memory and executive function.
- Slow Presentation Rate: Increase stimulus onset asynchrony (SOA) to 150-200 ms to accommodate slower processing speeds.
- Sensory Limitations (e.g., Visual Impairment):
- Enlarge Stimuli and Enhance Contrast: Ensure stimuli are sufficiently large and have high contrast against the background.
- Auditory Translation: Adapt the task for the auditory modality. Present a stream of sounds differing in two features (e.g., pitch and timbre) and ask participants to identify the features of a target sound.
Visual Short-Term Memory (VSTM) Binding Task
Original Protocol [64]:
- Objective: To assess the ability to bind features (e.g., shape and colour) in VSTM, a function particularly sensitive to Alzheimer's disease.
- Stimuli: Two arrays of three items each are presented briefly (200 ms) to the left and right of a fixation cross.
- Conditions:
- Shape Condition: Items are black shapes.
- Bind Condition: Items are shapes in different colours.
- Task: After a retention interval (900 ms), a test display appears. In the Shape condition, participants judge if the shapes are the same or if two are new. In the Bind condition, they judge if the shape-colour bindings are the same or if two shapes have swapped colours [64].
- Neural Measures: EEG can be used to analyse functional connectivity in frontal and occipital modules during encoding and maintenance periods.
Adaptations for Clinical Populations:
- Cognitive Limitations:
- Reduce Memory Set Size: Decrease the number of items per array from three to two or one to reduce VSTM load.
- Extend Encoding and Retention Time: Increase the study display duration from 200 ms to 500-1000 ms and lengthen the retention interval to reduce time pressure.
- Minimize Verbal Mediation: Use non-nameable shapes and non-primary colours to prevent verbal rehearsal, which may be variably affected in clinical populations [64].
- Sensory Limitations:
- Simplify Stimuli: Use highly distinct, simple shapes and highly discriminable colours to accommodate visual acuity or colour vision deficits.
- Spatial Cuing: Use arrows to direct attention to the relevant hemifield, reducing the visual search area [64].
Retro-Cue Paradigm in Visual Working Memory
Original Protocol [65]:
- Objective: To study how internal attention is directed within VWM to enhance performance.
- Stimuli: Participants remember a multi-feature array (e.g., items with colour and orientation). After the array disappears, a retro-cue is presented.
- Cue Types:
- Spatial Retro-cue: An arrow indicates the location of the relevant item.
- Feature Retro-cue: A feature (e.g., a colour patch) indicates which item's other feature (e.g., its orientation) will be tested.
- Task: After a variable cue-probe delay (50-650 ms), a probe appears, and participants make a same/different judgment. The retro-cue benefit (RCB) is the performance improvement in cued versus uncued trials [65].
Adaptations for Clinical Populations:
- Cognitive Limitations:
- Optimize Cue Type and Timing: Evidence suggests colour retro-cues produce benefits faster (at 50 ms delays) than spatial cues (200 ms) or orientation cues (500 ms) [65]. For populations with slowed processing, using colour cues and providing sufficient post-cue delay (>500 ms) can maximize the RCB.
- Explicit Task Instruction: Ensure participants are explicitly required to encode both features of an item, as orientation cues are only effective under this condition [65].
- Sensory Limitations:
- Enhance Cue Salience: Make retro-cues larger and higher contrast to ensure they are perceived.
- Tactile Adaptation: For visually impaired populations, the task could be adapted to a haptic version where objects vary in texture and shape.
Diagram 1: VWM Retro-Cue Task Workflow. The process from stimulus encoding to response, highlighting the critical stage of internal attention reallocation triggered by the retro-cue.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Materials and Tools for Temporal Binding Research
| Research Reagent / Tool | Function / Description | Example Use in Protocol |
|---|---|---|
| Non-nameable Shapes | Complex, hard-to-verbalize shapes prevent verbal rehearsal strategies, isolating visual memory processes. | Used in VSTM Binding Task to ensure pure assessment of visual, not verbal, memory [64]. |
| Non-primary Colours | Distinct, non-basic colours (e.g., mauve, teal) further minimize verbal labeling and rehearsal. | Paired with non-nameable shapes in the Bind condition of the VSTM task [64]. |
| E-Prime / PsychoPy | Software for precise stimulus presentation and response collection in cognitive psychology experiments. | To implement RSVP streams with millisecond accuracy and control response screen layout [62]. |
| EEG with High-Density Array | Electrophysiological recording to measure brain dynamics with high temporal resolution. | To analyse functional connectivity in frontal/occipital modules during VSTM binding [64] or ERP old/new effects in temporal memory [21]. |
| fMRI Scanner | To measure task-evoked and resting-state blood-oxygen-level-dependent (BOLD) signals. | To investigate hippocampal and entorhinal cortex representations of temporal context over long timescales [63]. |
| Modular Dirichlet Energy (MDE) | A novel graph signal processing technique to analyse transient functional network dynamics with high temporal precision. | Used with EEG to discern driving effects in brain modules (e.g., occipital 100-140 ms, frontal 140-180 ms) during VSTM tasks [64]. |
Data Presentation and Analysis of Key Findings
Summarizing quantitative data from foundational studies is critical for informing the design and interpretation of adapted protocols.
Table 3: Summary of Key Temporal Binding and Memory Findings
| Study Paradigm | Key Dependent Variable | Main Quantitative Finding | Clinical Interpretation |
|---|---|---|---|
| RSVP Temporal Binding [62] | Association between reported identity and colour | Significant within- and between-individual association: reporting a distractor's identity increased the likelihood of reporting its colour (p < 0.001). | Suggests a unified attentional episode; its delay or imprecision is a key target for clinical assessment. |
| Resting-State Temporal Drift [63] | Correlation of HPC/EC connectivity similarity with time interval | Negative correlation in both subjects (e.g., EC: r = -0.206, p < 0.001; HPC: r = -0.187, p < 0.001). aHPC showed stronger drift than pHPC. | The intrinsic "time tracker" is quantifiable; its disruption may be an early neural marker of pathology. |
| VSTM Binding & EEG [64] | Modular Dirichlet Energy (MDE) in Bind vs. Shape | In right-screen condition, Bind connectivity was less integrated than Shape in occipital (100-140 ms) and frontal (140-180 ms) modules. | Provides a precise temporal window (encoding period) to examine functional disconnection in clinical groups. |
| Retro-Cue Benefits [65] | Minimum effective cue-probe delay | Colour cues: 50 ms. Spatial cues: 200 ms. Orientation cues: 500 ms. | Informs cue selection and timing for populations with slowed cognitive processing to ensure measurable benefits. |
Integrated Workflow for Protocol Adaptation
Diagram 2: Clinical Protocol Adaptation Workflow. A systematic approach to modifying temporal binding and memory tasks for clinical populations, from initial assessment to validation.
Effectively adapting temporal binding protocols for clinical populations requires a principled approach grounded in the cognitive and neural mechanisms of feature binding and temporal context memory. Key strategies include simplifying stimuli and task demands, adjusting temporal parameters to accommodate slower processing speeds, and leveraging recognition-based responses to reduce cognitive load. Furthermore, integrating multimodal neural measures, such as EEG and fMRI, can provide sensitive biomarkers that complement behavioral data, especially in populations where performance is floor-bound by cognitive or sensory impairments. The protocols and adaptations outlined herein provide a robust foundation for researchers and drug development professionals to reliably assess episodic memory function in a wider range of clinical populations, thereby enhancing early detection and evaluation of therapeutic interventions.
Application Note: Core Stimulus Parameters for Temporal Binding
Optimizing stimulus parameters is fundamental to eliciting robust and interpretable neural and behavioral responses in temporal binding tasks for episodic memory research. The key parameters, their functional roles, and empirically-derived optimal values are summarized below.
Table 1: Key Stimulus Parameters for Temporal Binding Tasks
| Parameter | Functional Role in Temporal Binding | Empirical Finding / Recommended Value | Relevant Neural/Behavioral Readout |
|---|---|---|---|
| Phase Duration | Influences activation threshold and spatial specificity of neural response. | Shorter durations (e.g., 500 µs) elicit cortical activation at lower charge thresholds [66]. | Electrically Evoked Potentials (EEPs), Charge Threshold [66] |
| Longer durations (e.g., 1000-1500 µs) result in a more confined spread of cortical activation [66]. | Spatial Spread of Cortical Activation [66] | ||
| Stimulation Frequency | Affects neural adaptation and response fidelity to repetitive stimuli. | Responses are significantly attenuated at high frequencies (10-20 Hz) compared to low frequency (1 Hz) stimulation [66]. | Amplitude of Cortical EEPs [66] |
| Interphase Interval (IPI) | Can limit the extension of cortical responses, improving spatial resolution. | Inclusion of an IPI results in a more confined spread of cortical activation [66]. | Spatial Spread of Cortical Activation [66] |
| Auditory Probe Interval | Impacts signal-to-noise ratio and temporal resolution for evaluating mental workload. | Long, variable intervals (avg. 600 ms) are optimal for evaluating mental workload in detail using ERPs [67]. | Auditory N1/P2 ERP Components [67] |
Experimental Protocol: A Dynamic Emotional State Paradigm for Episodic Memory Structure
This protocol details a method to investigate how dynamic emotional states, elicited by music, shape the episodic structure of memory, providing a framework for temporal binding tasks [68].
Materials and Reagents
Table 2: Research Reagent Solutions for Musical Emotion Paradigm
| Item | Function in Protocol | Specification / Notes |
|---|---|---|
| Custom Musical Compositions | To elicit dynamic, fluctuating emotional states (valence and arousal) during encoding. | Professionally composed pieces conveying distinct emotional themes (e.g., joyous, anxious, sad, calm); Segmented into 30-40s emotional segments [68]. |
| Emotion Compass Tool | To collect continuous, moment-to-moment ratings of subjective emotional experience. | A software tool for tracking continuous valence and arousal ratings in real-time [68]. |
| Neutral Object Images | Used as the to-be-remembered stimuli for episodic memory tests. | Allows isolation of emotion's effect on memory structure without the confound of emotional item content [68]. |
| Event Segmentation Interface | To identify perceptual event boundaries in the music from an independent group of annotators. | Standard methodology where annotators press a button upon perceiving a meaningful change in the music [68]. |
Procedure
Day 1: Encoding Session
- Participant Setup: Seat the participant in a sound-attenuated room. Provide instructions for the Emotion Compass tool.
- Encoding Task: Present a series of custom musical compositions. Simultaneously, display sequences of neutral object images (e.g., one image every 2-4 seconds) on a computer screen.
- Continuous Emotion Tracking: While listening to the music and viewing the images, participants use the Emotion Compass to provide continuous ratings of their felt valence and arousal.
Day 2: Retrieval Session (24+ hours later)
- Item Recognition: Present participants with a mix of old (studied) and new (non-studied) object images. Ask them to identify whether each item is "old" or "new."
- Temporal Source Memory: For items correctly identified as "old," ask participants to indicate during which musical segment (e.g., first, second, or third) the item was originally encountered.
- Temporal Order Memory: Present pairs of old images and ask participants to judge which of the two appeared earlier in the study sequence.
- Subjective Distance Judgments: For the same pairs of images, ask participants to rate the subjective temporal distance between the items on a scale (e.g., "felt very close in time" to "felt very far apart in time").
Data Analysis
- Identify Emotional Boundaries: Perform change-point analysis on the continuous valence and arousal time series from the Emotion Compass to identify significant shifts in emotional state [68].
- Identify Musical Boundaries: Create a consensus perceptual boundary annotation from the independent event segmentation task [68].
- Analyze Memory Performance:
- Compare temporal order memory accuracy and subjective distance judgments for image pairs that cross an emotional/musical boundary versus pairs that reside within the same emotional/musical context.
- Analyze item recognition and temporal source memory for items that coincide with an emotional boundary versus those that do not.
Experimental Protocol: Optimizing Auditory Probes for Mental Workload Assessment
This protocol is adapted from research on evaluating mental workload using task-irrelevant auditory probes and is relevant for controlling cognitive load during complex episodic memory tasks [67].
Procedure
- Stimuli and Task:
- Primary Task: Participants perform a visually demanding cognitive task (e.g., a driving game at slow and fast speeds to impose low and high workload, respectively).
- Auditory Probes: Task-irrelevant auditory probe stimuli (e.g., simple tones) are presented through headphones during the primary task.
- Interval Manipulation: Present the auditory probes using different time interval conditions. The study manipulated [67]:
- Mean Interval Length: Long (600 ms) vs. Short (300 ms).
- Interval Variability: Variable (with five levels of jitter) vs. Fixed.
- EEG Recording: Continuous electroencephalogram (EEG) is recorded throughout the task to capture event-related potentials (ERPs) time-locked to the onset of the auditory probes.
Data Analysis
- ERP Analysis: Preprocess the EEG data and epoch around each auditory probe. Calculate the average amplitude for the N1 and P2 components.
- Workload Contrast: For each interval condition, contrast the N1 and P2 amplitudes between the low and high workload conditions.
- Temporal Resolution: Estimate the minimum data length required to obtain a statistically significant difference in ERP amplitude between workload conditions.
Visual Workflows
Figure 1. Experimental workflow for dynamic emotional state and memory study
Figure 2. Logical relationships of core optimization parameters
Integrating Computational Modeling with Behavioral Data for Enhanced Mechanistic Insight
Application Note: Computational Modeling in Temporal Binding Research
Temporal binding, the phenomenon whereby the time interval between a cause and its effect is perceived as shorter than the same interval separating two unrelated events, provides a crucial window into the cognitive mechanisms underlying episodic memory and agency perception [69]. This perceptual illusion represents a fundamental way in which causal cognition and temporal perception interact, with significant implications for understanding how humans construct and retrieve episodic memories that contain causal relationships [69] [70]. For researchers and drug development professionals, temporal binding tasks offer a sensitive behavioral paradigm for probing the cognitive and neural mechanisms that may be disrupted in various neurological and psychiatric conditions affecting memory.
The integration of computational modeling with behavioral data from temporal binding tasks enables researchers to move beyond descriptive accounts and develop quantitative, testable hypotheses about the algorithmic processes underlying this phenomenon [71]. By fitting models to experimental data, we can probe the specific computations the brain performs when binding events together in memory, identify neural correlates of these computational variables, and better understand how pharmacological interventions might alter these processes [71]. This approach is particularly valuable for investigating the component processes involved in episodic memory formation and retrieval, where temporal relationships between events constitute a fundamental organizational principle.
Key Findings from Temporal Binding Research
Recent research has demonstrated that temporal binding is present and developmentally constant from childhood into adulthood, with no significant changes in magnitude across age groups ranging from 6-7 years to adulthood [69]. This surprising developmental stability suggests that temporal binding reflects a fundamental cognitive mechanism that emerges early and remains consistent throughout development. The magnitude of the binding effect is significantly greater when participants themselves are the cause of an outcome compared to when they are mere spectators, indicating that intentional action modulates the strength of temporal binding [69].
Qualitative aspects of temporal binding data follow the principles of multisensory accounts, where actions and their effects are integrated in a statistically optimal manner [70]. However, quantitative evidence from comprehensive model evaluations indicates that temporal binding cannot be fully reduced to multisensory integration alone [70]. Rather, multisensory integration appears to be one of several component processes underlying temporal binding on an individual level, suggesting a more complex mechanistic architecture involving multiple cognitive systems.
Table 1: Key Experimental Findings in Temporal Binding Research
| Finding | Experimental Support | Implications for Episodic Memory |
|---|---|---|
| Developmental stability | No change in binding magnitude across ages 6-7 to adulthood [69] | Suggests core mechanism of temporal organization in memory is established early |
| Intentional action modulation | Greater binding for self-generated vs. observed actions [69] | Supports role of agency in structuring episodic memories |
| Multisensory components | Qualitative fit with multisensory integration models [70] | Indicates multiple systems contribute to temporal memory binding |
| Non-reduction to simple integration | Quantitative rejection of full multisensory account [70] | Points to additional cognitive processes in temporal binding |
Experimental Protocols
Interval Estimation Protocol for Temporal Binding
Apparatus and Setup
- Stimulus Presentation Computer: Standard PC running experimental software (e.g., PsychoPy, E-Prime, or custom JavaScript) with precise timing capabilities
- Response Input Device: Standard keyboard or button box with millisecond accuracy response recording
- Display Monitor: Standard LCD/LED monitor with 60Hz+ refresh rate
- Auditory Feedback System: Speakers or headphones for delivery of auditory stimuli when required
Participant Training Phase
- Train participants to identify four distinct time intervals using categorical labels (e.g., "very short," "short," "long," "very long")
- Present each interval duration multiple times in random order with appropriate labels
- Provide corrective feedback during training to ensure participants can reliably distinguish between the four interval categories
- Continue training until participants reach a predetermined accuracy criterion (e.g., 80% correct identification)
Experimental Procedure
Causal Condition:
- Participant presses a button that causes a rocket to launch on screen after a specific delay
- Participant estimates the time interval between their button press and the rocket launch by selecting one of the four trained interval categories
- Multiple trials with varying delays presented in random order
Non-Causal Predictive Condition:
- Participant observes a predictive signal (e.g., color change, symbol appearance) that indicates the rocket will launch after a delay
- Participant estimates the time interval between the predictive signal and the rocket launch using the same categorical response options
- Multiple trials with varying delays presented in random order
Control Condition (Optional):
- Participant observes two unrelated events (e.g., two sequential rocket launches)
- Participant estimates the time interval between these unrelated events
Table 2: Experimental Conditions and Parameters for Temporal Binding Tasks
| Condition | Initial Event | Outcome Event | Causal Relationship | Measured Interval |
|---|---|---|---|---|
| Causal | Self-generated button press | Rocket launch | Direct causal link | Action → outcome |
| Non-causal predictive | Predictive signal | Rocket launch | Predictive but not causal | Signal → outcome |
| Control | First rocket launch | Second rocket launch | No causal relationship | Event → event |
Data Collection Parameters
- Number of trials: 20-40 per condition (counterbalanced)
- Interval durations: 200ms, 400ms, 600ms, 800ms (adaptable based on pilot results)
- Inter-trial interval: 1000-2000ms (randomized)
- Total session duration: Approximately 30-45 minutes
Computational Modeling Protocol
Model Design Considerations
Computational modeling of temporal binding behavior requires careful consideration of several key aspects [71]:
Scientific Question Identification: Clearly define what cognitive process you are targeting (e.g., working memory contributions to temporal binding, Bayesian causal inference mechanisms)
Process Engagement: Ensure your experimental design actually engages the processes you intend to model through appropriate task parameters and conditions
Behavioral Signature Verification: Confirm that signatures of the targeted processes are evident from simple statistics of the behavioral data before proceeding with computational modeling
Model Fitting and Comparison Workflow
Model Simulation:
- Run candidate models with different parameter settings to generate simulated behavioral data
- Analyze simulated data using the same methods as real data to verify models can produce qualitative patterns matching empirical observations
Parameter Estimation:
- Find the set of parameter values that best account for real behavioral data for each model
- Use maximum likelihood or Bayesian estimation procedures appropriate for the specific model architecture
Model Comparison:
- Compute which of a set of possible models best describes the behavioral data
- Use appropriate comparison metrics (e.g., AIC, BIC, cross-validation) that account for model complexity
Latent Variable Inference:
- Use the best-fitting model to compute values of hidden variables not directly observable in behavioral data
- Relate these latent variables to neural measures (e.g., fMRI, EEG) or individual differences
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Materials and Computational Tools for Temporal Binding Research
| Item Category | Specific Examples | Function in Research | Implementation Notes |
|---|---|---|---|
| Experimental Software | PsychoPy, E-Prime, jsPsych | Precise stimulus presentation and response collection | Ensure millisecond timing accuracy; validate with external measurement devices |
| Response Collection Devices | Serial response boxes, gaming controllers, standard keyboards | Accurate recording of response times with minimal latency | Select based on required precision; serial devices typically offer ~1ms accuracy |
| Computational Modeling Frameworks | Stan, PyMC, TensorFlow Probability, MATLAB | Implementation and fitting of computational models | Choose based on model complexity and researcher expertise |
| Model Comparison Metrics | AIC, BIC, WAIC, Bayes factors | Quantitative comparison of different computational models | Use multiple metrics to assess robustness of conclusions |
| Data Visualization Tools | Matplotlib, ggplot2, Seaborn | Creation of publication-quality figures | Ensure accessibility compliance for color contrast [72] [73] |
| Statistical Analysis Platforms | R, Python (scipy, statsmodels), JASP | Statistical testing and data exploration | Implement reproducible analysis pipelines |
Integration with Episodic Memory Research
Neural Correlates of Temporal Binding
The frontoparietal and cingulate networks, which facilitate the retention of original memories, are likely involved in temporal binding processes that contribute to episodic memory formation [74]. These networks support effective conflict resolution during memory retrieval, which may be crucial for accurately binding causally related events in memory while maintaining their temporal structure [74].
Weak conflict control, coupled with intensified visual integration processing of interfering information, promotes the updating of original memories [74]. This suggests that temporal binding mechanisms might be modulated by similar processes during memory reconsolidation, potentially explaining how temporal relationships in episodic memories can be updated or distorted over time.
Methodological Considerations for Memory Research
When adapting temporal binding paradigms for episodic memory research, several key considerations emerge:
Encoding-Retrieval Design: Implement a three-phase design (encoding, interference/retrieval, testing) to examine how temporal binding affects long-term memory formation and retrieval [74]
Neuromodulation Approaches: Consider incorporating targeted neuromodulation techniques (e.g., tDCS) during critical memory phases to probe causal involvement of specific brain regions in temporal binding [74]
Multisensory Integration Assessment: Include measures of basic multisensory integration alongside temporal binding tasks to disentangle domain-general from specific temporal binding mechanisms [70]
Data Analysis and Visualization Protocols
Quantitative Data Management
For effective analysis of temporal binding data, implement the following procedures:
Data Tabulation:
- Create frequency tables for all categorical variables
- Ensure tables include clear titles, numbered sequentially (Table 1, Table 2, etc.)
- Present data in logical order (e.g., by size, importance, chronological sequence)
- Place percentages or averages to be compared as close as possible [75]
Class Interval Construction for Reaction Time Data:
- Calculate the range from lowest to highest value
- Divide this range into equal subranges (class intervals)
- Use between 6-16 classes for optimal balance between detail and conciseness
- Count frequencies for each class interval [75]
Visualization Standards
Histogram Creation:
Comparative Visualization:
- Use comparative histograms or bar charts when comparing conditions (e.g., causal vs. non-causal)
- Implement frequency polygons to emphasize distribution patterns when comparing multiple datasets
- For time trend data, employ line diagrams where class intervals represent temporal units [76]
Accessibility Compliance:
This comprehensive protocol provides researchers with the necessary methodological framework to integrate computational modeling with behavioral data from temporal binding tasks, offering enhanced mechanistic insight into the cognitive processes underlying episodic memory formation and retrieval.
Cross-Population Validation and Comparative Analysis of Temporal Binding Phenomena
Temporal Binding Window (TBW) alterations represent a significant transdiagnostic phenomenon in clinical neuroscience, offering a mechanistic bridge between basic sensory processing and higher-order cognitive deficits. This review synthesizes current evidence on TBW profiles in Obsessive-Compulsive Disorder (OCD), Mild Cognitive Impairment (MCI), and Addiction, framing these alterations within the context of episodic memory research. The TBW, defined as the temporal range within which disparate sensory inputs are perceptually fused into a unified event, serves as a critical foundation for constructing coherent episodic memories [77] [78]. In clinical populations, alterations in TBW width and plasticity may underlie fundamental disruptions in how temporal experiences are encoded, bound, and retrieved. This synthesis of multisensory integration (MSI) and memory frameworks provides novel insights for developing sensory-based biomarkers and interventions for neuropsychiatric disorders.
Clinical Profiles & Quantitative Data
TBW Alterations Across Clinical Populations
Table 1: TBW Characteristics Across Clinical Populations
| Clinical Population | TBW Profile | Key Findings | Neural Correlates | Relationship to Episodic Memory |
|---|---|---|---|---|
| Obsessive-Compulsive Disorder (OCD) | Wider pre-training TBW; abnormal tactile dominance; enhanced plasticity [77] [78] | • Larger visuo-tactile TBW pre-training• Tactile-leading stimulus dominance• Higher training gain (∼10-15% greater improvement) after perceptual training | • Altered prior weighting• Exaggerated reliance on real-time sensory input• Difficulties using past experiences | Disrupted formation of stable prior experiences, potentially affecting temporal context memory |
| Mild Cognitive Impairment (MCI) & Alzheimer's Pathology | Indirect evidence from bioimpedance & body composition [79] | • Reduced body cell mass/function in lower extremities (effect size: 0.26-0.51)• Altered cellular water distribution• Significant displacement on RXc graphs | • Segmental bioelectrical changes (↓ reactance/phase angle)• Association with cortical Aβ burden• Lower extremity neuromuscular degradation | Somatic markers of cognitive decline; potential peripheral proxy for central temporal processing deficits |
| Addiction | Striatal circuit dysfunction affecting temporal prediction [80] | • Maladaptive corticostriatal neuroadaptations• Altered dopamine-dependent plasticity• Hijacked reward prediction error signals | • Dorsal/ventral striatal pathway alterations• Prefrontal-striatal circuit synaptic potentiation• Mesolimbic dopamine system dysregulation | Compromised temporal prediction mechanisms crucial for encoding reward contingencies in episodic memory |
Table 2: Experimental Data and Effect Sizes
| Parameter | OCD (vs. Healthy Controls) | MCI/AD (vs. Cognitively Normal) | Addiction Models |
|---|---|---|---|
| Sample Sizes | 31 OCD, 34 controls [77] | 365 CN, 123 MCI due to AD, 30 AD dementia [79] | Animal models (various) [80] |
| Primary Effect Sizes | Large TBW difference pre-training; >10% training gain | Effect sizes 0.26-0.51 for lower extremity parameters [79] | Not quantified in human TBW studies |
| Modality Specificity | Visuo-tactile dominance [77] [78] | Segmental bioelectrical (lower extremities) [79] | Cross-modal reward prediction errors |
| Plasticity Profile | Enhanced training responsiveness | Progressive deterioration | Long-lasting synaptic plasticity |
Experimental Protocols
Visuo-Tactile Simultaneity Judgment Task for OCD
Application: Assessing and modulating TBW width in OCD populations [77] [78].
Materials:
- Computer with precision timing software
- Visual display unit
- Tactile stimulator (e.g., tactor)
- Response input device
Procedure:
- Pre-training Baseline Assessment:
- Present visuo-tactile stimulus pairs with varying stimulus onset asynchronies (SOAs)
- Use SOAs ranging from -500ms (tactile-first) to +500ms (visual-first)
- Participants perform simultaneity judgments ("simultaneous" vs. "sequential")
- Calculate TBW width from psychometric function (range of SOAs perceived as simultaneous)
Perceptual Training Phase:
- Administer repetitive simultaneity judgments with adaptive SOA narrowing
- Provide trial-by-trial feedback on performance accuracy
- Utilize staircase procedures to progressively narrow SOAs around perceptual threshold
- Conduct multiple sessions (typically 3-5) over consecutive days
Post-training Assessment:
- Re-administer identical simultaneity judgment task from baseline
- Calculate TBW width and compare to pre-training measures
- Derive training gain scores as index of TBW plasticity
Data Analysis:
- Fit Gaussian functions to simultaneity judgment data
- Define TBW width as ±1 SD from the mean of the fitted function
- Compare pre-post TBW changes between groups (OCD vs. controls)
Libet's Clock Protocol for Temporal Binding
Application: Measuring intentional binding and sense of agency across clinical populations [53] [56] [81].
Materials:
- Libet's clock display (clock face with rotating dot, 2560ms/rotation)
- Response button
- Auditory tone generator (1000Hz)
- Precision timing software (e.g., Labclock Web) [53] [56]
Procedure:
- Setup and Instructions:
- Present clock with rotating dot (one full rotation as familiarization)
- Instruct participants to press button voluntarily during second rotation
- For operant trials, tone follows action after specified delay (250ms)
- Participants estimate time of action or tone using clock position
Trial Types (Blocked Design):
- Agency Action Trials: Key press triggers tone after 250ms delay; report key press time
- Agency Outcome Trials: Key press triggers tone after 250ms delay; report tone time
- Baseline-Action Trials: Key press only; report key press time
- Baseline-Outcome Trials: Tone only; report tone time [56]
Temporal Binding Calculation:
- Compute shift in perceived time of action toward outcome (action binding)
- Compute shift in perceived time of outcome toward action (outcome binding)
- Compare operant vs. baseline conditions for binding effects
Clinical Modifications:
- For OCD: Incorporate rule-breaking conditions to assess cognitive conflict [81]
- For addiction: Include drug-related cues to assess stimulus-specific binding
- For MCI: Simplify task (reduce trials) and extend response windows
Segmental Bioimpedance Analysis for MCI/AD
Application: Assessing segmental body composition changes as peripheral proxies of central pathology [79].
Materials:
- Multifrequency bioimpedance analyzer (e.g., InBody S10)
- Standard anthropometric measurement tools
- Examination table for supine positioning
Procedure:
- Participant Preparation:
- Ensure no acute hydration imbalances
- Measure height, weight, and BMI
- Position supine with limbs abducted from body
Segmental Measurement:
- Apply tetrapolar 8-point tactile electrodes
- Measure impedance at six frequencies (1, 5, 50, 250, 500, 1000 kHz)
- Record reactance and phase angle at three frequencies (5, 50, 250 kHz)
- Obtain segmental values for arms, legs, and trunk
Data Processing:
- Calculate extracellular water to total body water ratio (ECW/TBW)
- Compute phase angle and body cell mass estimates
- Generate RXc graphs for vector analysis
Analysis Approach:
- Compare segmental parameters between CN, MCI, and AD dementia groups
- Correlate bioelectrical measures with cognitive test scores
- Analyze vector displacement on confidence ellipses
Methodological Visualization
Experimental Workflow for Clinical TBW Assessment
Neural Circuits in TBW Pathology
The Scientist's Toolkit
Table 3: Essential Research Reagents and Solutions
| Tool/Reagent | Primary Application | Function/Significance | Example Specifications |
|---|---|---|---|
| Labclock Web Software [53] | Temporal binding assessment | Open-source HTML5 implementation of Libet's clock paradigm; enables precise timing measurement for action-outcome binding | • 2560ms rotation period• Customizable delays (100-700ms)• Online administration capability |
| Multifrequency Bioimpedance Analyzer [79] | Segmental body composition analysis in MCI/AD | Measures segmental impedance, reactance, and phase angle as proxies for cellular integrity and hydration status | • InBody S10 system• 6 frequencies (1-1000 kHz)• 8-point tactile electrode system |
| Visuo-Tactile Stimulation Apparatus [77] [78] | Simultaneity judgment tasks | Presents precisely timed visual and tactile stimuli for TBW boundary assessment | • LCD visual display• Tactile stimulators (vibratory)• Sub-millisecond timing precision |
| fMRI-Compatible Response Devices [22] | Neural correlates of temporal binding | Enables measurement of BOLD response during serial encoding and recall paradigms | • Fiber-optic response pads• MRI-compatible buttons• Precision timing synchronization |
| Striatal Pathway-Specific Viral Vectors [80] | Addiction circuit manipulation | Enables optogenetic/chemogenetic dissection of striatal circuits involved in temporal prediction | • D1R/D2R-Cre dependent constructs• Pathway-specific tracers• Cell-type specific promoters |
Discussion and Integration with Episodic Memory Research
The TBW alterations observed across OCD, MCI, and addiction profiles reveal distinct but complementary disruptions in the temporal architecture of experience. In OCD, the widened TBW and paradoxical enhanced plasticity [77] [78] suggest a fundamental imbalance in how prior experiences are weighted against real-time sensory evidence. This aligns with computational accounts of OCD as a disorder of inference and uncertainty, where compromised priors disrupt the formation of stable temporal representations necessary for episodic memory.
The MCI and Alzheimer's pathology findings, while not directly measuring TBW, reveal peripheral manifestations of temporal processing degradation through segmental bioimpedance alterations [79]. The particular vulnerability of lower extremity parameters suggests a somatotopic mapping of cognitive decline, potentially reflecting the hierarchical organization of temporal processing networks. This peripheral profiling offers a complementary approach to understanding central temporal integration deficits in neurodegenerative disorders.
In addiction profiles, the striatal circuit dysregulation [80] points to a different mechanism of temporal binding disruption—one centered on reward prediction error signaling and temporal contiguity assessment. The maladaptive prioritization of drug-related temporal associations may directly compete with and disrupt the formation of adaptive episodic memories, creating a vicious cycle where substance-related temporal predictions dominate cognitive resources.
Across all three populations, the connection to episodic memory emerges through shared mechanisms of temporal context binding [22]. The hippocampus and related medial temporal lobe structures, which show robust involvement in serial encoding and event segmentation, likely serve as the critical nexus where TBW alterations translate into episodic memory deficits. Future research should directly examine how TBW width correlates with specific aspects of temporal memory, particularly the binding of items to their temporal context and the segmentation of continuous experience into discrete events.
TBW assessment provides a valuable transdiagnostic framework for understanding clinical disruptions in episodic memory. The distinct profiles observed in OCD (wider TBW, enhanced plasticity), MCI/AD (indirect bioimpedance markers), and addiction (striatal prediction errors) highlight the multifaceted nature of temporal processing across neuropsychiatric spectra. Methodological innovations in simultaneity judgment tasks, Libet's clock paradigms, and segmental bioelectrical assessment offer robust tools for quantifying these alterations.
Future research should prioritize:
- Longitudinal designs tracking TBW changes across disease progression
- Multimodal approaches combining behavioral TBW measures with neuroimaging and electrophysiology
- Interventional studies examining whether TBW normalization correlates with cognitive improvement
- Cross-species paradigms [82] enabling circuit-level manipulation of temporal binding mechanisms
By bridging multisensory integration, temporal binding, and episodic memory research, this integrated approach offers promising pathways for developing novel biomarkers and sensory-based interventions for neuropsychiatric disorders.
Application Notes
Emotional episodic memory is a cornerstone of human experience, and its modulation by psychoactive drugs has significant implications for both therapeutic development and understanding substance misuse. A core aspect of episodic memory is temporal binding—the process by which the brain links the diverse elements of an experience (sensory data, context, emotional valence) into a coherent, unified memory trace over time. Different drug classes distinctly modulate the neural systems responsible for this binding process, particularly during the encoding, consolidation, and retrieval of emotional memories. Understanding these differential mechanisms is critical for designing targeted interventions for affective and trauma-related disorders and for interpreting the cognitive effects of recreational drug use within a neurocognitive framework of episodic memory.
Key Neurobiological Systems Involved
The modulation of emotional memory involves a complex interplay of brain networks:
- Medial Temporal Lobe (MTL) and Hippocampus: Central to the formation and temporal binding of episodic memories [83]. The hippocampus is crucial for integrating information into cohesive engrams.
- Amygdala: Strongly interacts with the hippocampus to modulate the emotional strength of memories, particularly for arousing content [83].
- Prefrontal Cortex (PFC): Regions like the dorsolateral PFC (DLPFC) and medial PFC (mPFC) support cognitive control and organization during memory encoding and retrieval [83].
- Parietal Cortex: Involved in directing attentional resources toward internal memory representations, facilitating their maintenance and retrieval [83].
- Visual and Sensory Integration Cortex: The Occipital Fusiform Gyrus (OFG) and related areas are pivotal for integrating new sensory information, which can promote the updating or distortion of existing memories [74].
Drugs exert their effects by modulating the neurotransmitter systems that regulate the activity and functional connectivity within this network.
Table 1: Comparative Effects of Drug Classes on Emotional Episodic Memory Stages
| Drug Class | Primary Neurotransmitter Action | Effect on Encoding of Emotional vs. Neutral Memory | Effect on Consolidation of Emotional vs. Neutral Memory | Effect on Retrieval of Emotional vs. Neutral Memory | Key Brain Regions/Networks Modulated |
|---|---|---|---|---|---|
| GABAA Sedatives (e.g., Alcohol, Benzodiazepines) | Positive allosteric modulation of GABAA receptors | Preferentially impairs emotional memory [58] | Preferentially enhances emotional memory (effect may reverse with long delay) [58] | May distort memory, potentially with selectivity for emotional (positive) memory [58] | Frontoparietal network; Hippocampus-Amygdala interaction [83] |
| THC (Cannabis) | CB1 receptor agonist | Preferentially impairs emotional memory [58] | Information Not Specified | Distorts memory, with potentially greater selectivity for emotional (positive) memory [58] | Hippocampus; Prefrontal Cortex [58] |
| Stimulants (e.g., Dextroamphetamine) | Dopamine & Norepinephrine transport inhibitors | Can preferentially enhance emotional memory (less consistent) [58] | Information Not Specified | Distorts memory, with potentially greater selectivity for emotional memory [58] | Frontoparietal control networks; Cingulo-opercular network [74] [83] |
| MDMA | Serotonin, Dopamine, Norepinephrine transporter inhibitor; 5-HT2A receptor agonist | Can both impair and enhance emotional memory (effects vary) [58] | Information Not Specified | Distorts memory, with potentially greater selectivity for emotional (positive) memory [58] | Networks involving 5-HT2A receptor activation [58] |
| Ketamine | NMDA receptor antagonist | Preferentially impairs emotional memory [58] | Information Not Specified | Little impact on retrieval (when isolated) [58] | Networks involving NMDA receptor function [58] |
Experimental Protocols
General Framework for Temporal Binding and Emotional Memory Assessment
The following protocol outlines a three-phase design suitable for investigating drug effects on emotional episodic memory, incorporating elements of temporal binding and post-retrieval updating [74].
3.1.1. Protocol: Three-Phase Emotional Memory and Updating Task
Objective: To assess the effect of a drug intervention on the encoding, consolidation, and retrieval-updating of emotional memories within a temporal context.
Design: A three-day, within-subjects or between-subjects design.
- Day 1 (Encoding): Participants encode neutral object images superimposed on emotionally negative or neutral background scenes. This creates context-dependent episodic memories.
- Day 2 (Interference/Reactivation): Under the influence of the drug or placebo, participants' memories are reactivated. They are presented with some of the original scenes but now paired with new, conflicting neutral objects (interference condition). Other conditions include mere relearning (original pairings) and no-retrieval controls.
- Day 3 (Final Test): Memory for the original object-scene associations is tested to determine the degree of memory preservation (correct original memory) versus updating (incorrectly recalling the new object from Day 2) [74].
Key Dependent Variables:
- Accuracy of original memory (Preservation).
- Rate of incorrect updating with new information.
- fMRI or EEG correlates during the interference phase (e.g., activity in DLPFC, dACC, IPL for preservation; OFG for updating) [74].
Table 2: Research Reagent Solutions for Emotional Memory Protocols
| Item Name | Function/Brief Explanation | Example Application in Protocol |
|---|---|---|
| Standardized Affective Picture Sets | Provides validated visual stimuli with normative ratings of emotional valence and arousal. | Used as emotional and neutral background scenes during the encoding phase to provide the emotional context. |
| Neutral Object Image Library | A set of pictographic or photographic images of common objects that are emotionally neutral. | Used as the core item to be remembered, independent of its inherent emotional content. |
| fMRI-Compatible Presentation System | Software and hardware for precise stimulus timing and synchronization with fMRI scanner pulses. | Critical for presenting the memory task and collecting neural data during the interference phase on Day 2 [74]. |
| High-Definition tDCS System | Non-invasive brain stimulation device for modulating cortical excitability. | Can be used to target the visual cortex during reactivation to test causal involvement in memory updating [74]. |
| Continuous Emotion Tracking Tool | Software for collecting moment-to-moment ratings of valence and arousal (e.g., Emotion Compass). | Can be used during encoding or drug onset to track dynamic emotional states and identify emotional event boundaries that influence memory [68]. |
| Pharmacological Agent & Matched Placebo | The drug under investigation and an indistinguishable control substance. | Administered in a double-blind, randomized fashion before the target memory process (encoding, consolidation, or retrieval). |
Protocol for Isulating Drug Effects on Memory Consolidation
Objective: To specifically investigate the effect of a drug on the post-encoding stabilization (consolidation) of emotional memories.
Design:
- Encoding Phase (Drug-Free): Participants encode a series of emotional and neutral stimuli (e.g., images, words).
- Immediate Post-Encoding Drug Administration: Immediately after encoding, participants are administered the drug or placebo. Using a drug with rapid absorption is critical to ensure it is active during the cellular consolidation window [58].
- Retrieval Phase (Drug-Free): After a delay (e.g., 24 hours or longer, to ensure the drug has been fully metabolized), memory for the encoded stimuli is tested. This isolates the drug's effect to the consolidation phase, separate from encoding or retrieval.
Key Dependent Variables:
- Difference in recall or recognition accuracy for emotional items versus neutral items between the drug and placebo conditions.
Visualization of Mechanisms and Workflows
Drug Modulation of Memory Pathways
Diagram 1: Drug Modulation of Memory Pathways
Emotional Memory Experiment Workflow
Diagram 2: Emotional Memory Experiment Workflow
In the search for accessible and non-invasive biomarkers for neurodegenerative conditions, saccadic eye movements have emerged as a promising tool for detecting subtle cognitive decline. This application note details the use of saccadic accuracy and related oculometrics to differentiate Mild Cognitive Impairment (MCI), a potential prodromal stage of Alzheimer's disease dementia (ADD), from normal aging. This field is situated within the broader context of episodic memory research, which posits that the hippocampus provides a spatiotemporal framework for binding event details [18]. The integrity of this binding process, crucial for forming coherent episodic memories, is reflected in the executive control required to perform volitional saccadic tasks, thereby linking eye movement metrics to core mnemonic functions [84].
Meta-analyses and recent studies consistently reveal specific alterations in saccadic performance in MCI and ADD populations. The following tables summarize key oculometric parameters that serve as effective differentiators.
Table 1: Key Saccadic Parameters Differentiating MCI, ADD, and Healthy Controls (HC)
| Participant Group | Prosaccade Latency (ms) | Antisaccade Latency (ms) | Antisaccade Error Rate (%) | Critical Findings |
|---|---|---|---|---|
| Healthy Controls (HC) | ~220 [85] | ~343 [85] | ~0.7 [85] | Baseline for typical aging [85]. |
| Mild Cognitive Impairment (MCI) | Significantly longer than HC [86] | Significantly longer than HC [86] | Significantly higher than HC [86] | Differentiates from HC and FCD; impaired inhibitory control [87] [86]. |
| Alzheimer's Disease Dementia (ADD) | Longer than MCI [86] | Longer than MCI [86] | Higher than MCI [86] | Greater deficits in latency and error rate than MCI [86]. |
| Functional Cognitive Disorder (FCD) | Differs from HC [87] | Does not differ from HC [87] | Does not differ from HC [87] | Distinct profile: prosaccade deficits but intact antisaccade performance [87]. |
Table 2: Effect Paradigms in Saccadic Tasks
| Effect Paradigm | Definition | Utility in Differentiation |
|---|---|---|
| Gap Effect | Latency reduction when central fixation disappears before peripheral target appears. | The magnitude of the latency gap effect varies among HC, MCI, and ADD, aiding distinction [86]. |
| Anti-Effect | Difference in performance (latency, error rate) between prosaccade and antisaccade tasks. | Effectively distinguishes patients with ADD and MCI from controls, with antisaccade paradigms being more effective than prosaccades [86]. |
Experimental Protocols for Saccadic Assessment
The following are detailed protocols for core saccadic tasks, which can be implemented using eye-tracking systems in laboratory settings.
Protocol for the Prosaccade Task
The prosaccade task measures the integrity of basic, reflexive orienting systems [87].
- Objective: To assess the speed and accuracy of visually guided, reflexive saccades.
- Stimuli & Setup: Participants sit facing a display screen (e.g., monitor or VR headset). A central fixation point (e.g., a cross) and peripheral targets (e.g., a dot) are used.
- Procedure:
- The trial begins with the participant fixating on a central point.
- After a random interval (e.g., 1000-2000 ms), the central point extinguishes, and a target simultaneously appears at a predetermined peripheral location (e.g., 10° to the left or right).
- The participant is instructed to look at the peripheral target as quickly and accurately as possible.
- The target remains visible for a set duration (e.g., 1500 ms) before returning to the central fixation for the next trial.
- A typical block consists of ~120 trials with randomized target locations [85].
- Data Analysis:
- Latency: Mean and standard deviation of reaction time (from target onset to saccade initiation) [87].
- Accuracy: Amplitude or endpoint error of the primary saccade towards the target.
Protocol for the Antisaccade Task
The antisaccade task is a gold standard for assessing cognitive control, response inhibition, and working memory [87] [86].
- Objective: To assess the ability to inhibit a reflexive saccade and generate a volitional saccade to the mirror-opposite location.
- Stimuli & Setup: Identical to the prosaccade setup.
- Procedure:
- The participant fixates on a central point.
- After a random interval, the central point disappears, and a peripheral "distractor" stimulus appears (e.g., 10° to the left).
- The participant is instructed to actively inhibit looking at the distractor and instead make a saccade to the mirror-opposite location (e.g., 10° to the right) as quickly and accurately as possible.
- The distractor is typically displayed briefly (e.g., 100-200 ms) to prevent visual capture.
- A typical block consists of ~120 trials [85].
- Data Analysis:
- Error Rate: The proportion of trials where the first saccade is directed toward the distractor instead of the correct opposite location [87] [86].
- Latency: Mean and standard deviation of correct antisaccade reaction times [87].
- Correct Antisaccade Gain: The amplitude of correct antisaccades relative to the target location.
Diagram 1: Antisaccade task cognitive process.
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Materials and Equipment for Saccadic Research
| Item | Function/Description | Example Specifications |
|---|---|---|
| Video-Based Eye Tracker | Non-invasive measurement of eye position and pupil data using infrared cameras. | Sampling rate: ≥ 250 Hz (≥1000 Hz for high-resolution analysis) [85] [88]. Spatial resolution: 0.25°–0.5° [85]. |
| Stimulus Presentation System | Displays visual paradigms and records behavioral responses. | A computer monitor or a Head-Mounted Display (HMD) with integrated eye tracking (e.g., VIVE Pro Eye) for immersive control [85]. |
| Saccade Detection Algorithm | Processes raw gaze data to identify and classify saccades, fixations, and other ocular events. | Velocity-threshold algorithm (e.g., I-VT filter) [89]. Custom algorithms (e.g., in MATLAB/Python) to refine detection and unify fragmented saccades [89] [85]. |
| Analysis Software | Platform for statistical analysis and visualization of oculometric parameters. | MATLAB, R, or Python with custom scripts to calculate latency, velocity, amplitude, and error rates. |
Diagram 2: Saccadic assessment system workflow.
Integration with Episodic Memory Research
The connection between saccadic control and episodic memory is grounded in shared neural substrates, particularly the hippocampal formation and prefrontal cortex.
- Spatiotemporal Binding and the Hippocampus: Episodic memory requires the binding of "what," "where," and "when" information into a coherent trace. The hippocampus is theorized to provide a spatiotemporal framework for this binding process [18]. Successful memory formation relies on integrating information across spatial and temporal gaps, a function dependent on the hippocampus [84].
- Executive Control and Mnemonic Function: The antisaccade task directly probes response inhibition and working memory—executive functions that are also critical for controlling the encoding and retrieval of episodic memories [86]. The frontal-hippocampal network supports both volitional saccade generation and relational memory binding [18]. Deficits in antisaccade performance in MCI thus reflect a breakdown in this network that also underpins episodic memory impairment.
- A Non-Invasive Proxy: Given that eye movement parameters provide an indirect window into the integrity of these shared neural circuits, saccadic metrics can serve as a behavioral proxy for the efficacy of spatiotemporal binding processes that are more difficult to assess directly in clinical settings [87] [86].
Application Notes
Theoretical Foundations and Computational Principles
Computational models of memory provide a foundational framework for understanding the mechanisms underlying episodic recall, particularly the phenomenon of temporal binding—the process by which the brain links discrete events across time to form coherent episodic sequences. These models conceptualize memory not as a static store but as a dynamic system where items are associated with an evolving 'context' signal. During recall, items subsequently compete with one another based on these associations [90]. This context signal is crucial for explaining fundamental dynamics of episodic memory, such as contiguity effects (the tendency to recall items that were proximal in time) and recency effects (the superior recall for the most recent items) [22] [90].
The core computational challenge is modeling how serial order is maintained. Two major theories have been debated: associative chaining, which posits direct inter-item associations, and positional coding, which advocates for item-position associations. Recent behavioral evidence, including the finding that shuffling item positions while keeping relative order intact does not interfere with serial recall, lends stronger support to associative chaining theories. This suggests serial recall involves the mnemonic binding of one item to the next across a temporal gap [22]. Computational models implementing these principles, such as the Temporal Context Model (TCM), use a gradually shifting context representation to explain how temporal associations are formed and retrieved [90].
A critical prediction of these models is that contextual stability modulates the binding process. Behaviorally, serial recall is greater for information encountered within the same event compared to across event boundaries. Computationally, this occurs because an event boundary induces a more rapid shift or reset in the underlying context signal, making it more difficult to form associative links across the boundary. The models predict that bridging these temporal gaps requires additional cognitive computation, potentially mediated by prefrontal-hippocampal interactions [22].
Key Model Predictions for Temporal Binding
Computational models generate several testable predictions regarding the neural and behavioral correlates of temporal binding in episodic recall:
- Prediction 1: Hippocampal-Mediated Context Binding. Models predict the hippocampus is critical for associating items with an evolving temporal context. This is reflected in increasing neural pattern similarity within the same sequence over repetitions, a signature of sequence learning [22] [90].
- Prediction 2: Prefrontal Cortex in Active Retrieval and Integration. The models predict that the ventrolateral prefrontal cortex (vlPFC), particularly when binding across event boundaries, actively retrieves or refreshes pre-boundary representations to integrate consecutive items separated by a contextual shift [22].
- Prediction 3: Distinct Neural Profiles for Preservation vs. Updating. Models of memory reconsolidation and updating predict that the retention of original memories is associated with strong engagement of conflict control networks (e.g., frontoparietal and cingulo-opercular networks). In contrast, the successful updating of memories with new information is associated with intensified processing in sensory integration areas, such as the Occipital Fusiform Gyrus (OFG) [74].
- Prediction 4: Functional Connectivity Signatures. Beyond univariate activation, models predict that successful within-event binding is supported by increased functional connectivity between the hippocampus and ventromedial PFC, highlighting how different neural circuits support binding under different contextual conditions [22].
Table 1: Summary of Key Computational Model Predictions and Their Correlates
| Prediction | Computational Basis | Neural Correlate | Behavioral Manifestation |
|---|---|---|---|
| Contextual Binding | Gradually shifting context signal [90] | Hippocampus; Pattern similarity [22] | Contiguity Effect; Serial Recall [22] |
| Cross-Boundary Integration | Context reset at event boundaries [22] | vlPFC activation [22] | Reduced recall across vs. within events [22] |
| Memory Preservation | Effective conflict resolution [74] | Frontoparietal & Cingulo-opercular networks [74] | Accurate recall of original memory [74] |
| Memory Updating | New sensory integration [74] | Elevated Occipital Fusiform Gyrus (OFG) activity [74] | Incorporation of interfering information [74] |
Experimental Protocols
fMRI Protocol: Investigating Neural Mechanisms of Temporal Binding
This protocol is designed to test key model predictions on the neural substrates of temporal binding within and across event boundaries in humans [22] [74].
Experimental Design and Paradigm
- Participants: 25-30 right-handed native speakers, aged 18-28, screened for neurological conditions.
- Stimuli: Trial-unique images of celebrity faces and nameable objects.
- Procedure:
- Encoding Phase: Participants are presented with a series of 25 images in a pseudo-randomized sequence. The context (stimulus category and task) changes semi-predictably to create "event boundaries."
- Each image is displayed for 2 seconds, followed by a 2-second category-specific judgment task (e.g., likability for faces, commonality for objects).
- The Inter-Trial Interval (ITI) is jittered (4, 6, or 8 seconds) to orthogonalize the hemodynamic response.
- Distractor Task: A 45-second arrow discrimination task follows encoding to prevent rehearsal.
- Test Phase: Conducted outside the scanner after a delay.
- Serial Recall Test: Participants are given up to 90 seconds to recall the list of images in the exact order of presentation.
- Recency Discrimination Test (optional): Participants are shown 12 image pairs and indicate which image was presented more recently.
- Encoding Phase: Participants are presented with a series of 25 images in a pseudo-randomized sequence. The context (stimulus category and task) changes semi-predictably to create "event boundaries."
fMRI Data Acquisition and Analysis
- Acquisition Parameters: Use a 3T MRI scanner. Acquire T2*-weighted echoplanar imaging (EPI) sequences for BOLD contrast (e.g., TR=2000ms, TE=30ms, voxel size=3x3x3mm).
- Preprocessing: Standard pipeline including slice-time correction, realignment, normalization to MNI space, and smoothing with a Gaussian kernel (e.g., 8mm FWHM).
- Statistical Analysis:
- Univariate Analysis: Construct a general linear model (GLM) with regressors for:
- Item encoding, categorized by subsequent memory performance (Serially Recalled vs. Not Recalled) and event structure (Within-Event vs. Across-Boundary).
- Category judgment tasks.
- Functional Connectivity Analysis: Apply psychophysiological interaction (PPI) analysis to examine context-dependent changes in connectivity between the hippocampus and prefrontal regions (vmPFC, vlPFC) during the encoding of subsequently serially recalled items.
- Univariate Analysis: Construct a general linear model (GLM) with regressors for:
Rodent Behavioral Protocol: Temporal Order Memory Task
This protocol adapts the core principles of human temporal binding research for preclinical models, allowing for the testing of novel pharmacological agents and invasive manipulations [34].
Behavioral Apparatus and Materials
- Apparatus: A rectangular open field (e.g., 60cm x 40cm) made of white Plexiglas, placed in a room with distinct distal spatial cues.
- Objects: Use multiple copies of distinct, non-naturalistic objects (e.g., glass pyramids, metal cubes) that are too heavy for the animal to displace.
- Tracking: An overhead camera and video tracking software (e.g., EthoVision XT) to record the animal's position, head direction, and object interactions.
Behavioral Procedure (Sample Trial)
- Habituation: The rodent is allowed to freely explore the empty apparatus for 5-10 minutes on at least two consecutive days.
- Encoding Phase:
- Sample 1: The rodent is placed in the arena containing two identical copies of Object A (e.g., two pyramids) for a fixed period (e.g., 5 minutes).
- Delay: The rodent is returned to its home cage for a fixed delay (e.g., 1 hour). The delay interval can be manipulated as an independent variable.
- Sample 2: The rodent is placed in the arena containing two identical copies of a novel Object B (e.g., two cubes) for 5 minutes.
- Test Phase:
- After a retention delay (e.g., 1 hour), the rodent is returned to the arena containing three objects: a copy of Object A, a copy of Object B, and a novel Object C.
- The animal's exploration (sniffing, touching, with head oriented towards the object within <2cm) of each object is recorded for 5 minutes.
Data Analysis and Quantification
- Key Dependent Variable: The Temporal Memory Index.
- Calculation:
(Time with Oldest Object - Time with Middle Object) / Total Time with All Objects - A positive index indicates the animal spent more time with the object encountered farthest back in time (Object A), demonstrating successful temporal order memory.
- Calculation:
- Control Measures: Total object exploration time to ensure equal engagement. Discrimination between the novel Object C and the familiar Objects A and B to control for general novelty preference.
tDCS Intervention Protocol: Modifying Memory Updating
This protocol leverages model predictions about the role of sensory cortex in memory updating, offering a non-pharmacological approach to modulate memory processes [74].
Experimental Design
- Design: Double-blind, sham-controlled, between-subjects or within-subjects design.
- Groups: Participants are randomly assigned to either Active tDCS or Sham tDCS stimulation groups.
Procedure (Three-Phase Design)
- Day 1: Memory Encoding: Participants encode a set of original memories (e.g., object-location associations).
- Day 2: Interference under tDCS:
- Stimulation Setup: Apply high-definition tDCS with the anode positioned over the visual cortex (e.g., Oz based on the 10-20 EEG system) and the cathode over a contralateral supraorbital region.
- Stimulation Parameters:
- Active tDCS: 20-minute stimulation at 1.5 mA, with a 30-second ramp up/down.
- Sham tDCS: Identical setup and initial ramp-up/down, but no sustained current.
- Interference Task: During stimulation, participants are exposed to interfering information that conflicts with the original memories encoded on Day 1 (e.g., the same objects in new locations).
- Day 3: Final Memory Test: Participants are tested on both the original (Day 1) and the interfering (Day 2) information to assess memory preservation vs. updating.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Materials and Reagents for Temporal Binding Research
| Item / Reagent | Function / Application | Example Use in Protocol |
|---|---|---|
| Celebrity Face & Object Image Sets | Standardized stimuli for human memory tasks. | fMRI encoding paradigm; creates distinct "what" content [22]. |
| Video Tracking Software (e.g., EthoVision XT) | Automated recording and analysis of rodent behavior. | Quantifying object exploration and locomotion in rodent temporal order task [34]. |
| High-Definition tDCS System | Non-invasive neuromodulation of cortical activity. | Applying targeted stimulation to the visual cortex during memory interference to probe updating mechanisms [74]. |
| Analysis Libraries (SPM, FSL, AFNI) | Statistical analysis and modeling of fMRI data. | Performing univariate and functional connectivity (PPI) analyses to identify neural correlates of binding [22] [74]. |
| Computational Modeling Frameworks (e.g., TCM, CDP) | Formalizing theories and generating quantitative predictions. | Simulating recall dynamics and contiguity effects to test theories of associative chaining vs. positional coding [22] [90]. |
| Distinct, Non-Naturalistic Objects | Stimuli for rodent novel object and temporal order memory tasks. | Serving as memorable "what" content in rodent behavioral protocols (Objects A, B, C) [34]. |
Item-in-context memory, a fundamental component of episodic memory, relies on the hippocampal formation to bind items with the contextual details of their occurrence. This Application Note synthesizes current research to establish a validated cross-species framework for translating rodent models of item-in-context memory to human applications, with emphasis on pharmacological testing and cognitive assessment. We provide detailed protocols for behavioral tasks, neural analysis, and computational modeling that enable direct comparison across species, addressing the critical translational gap in therapeutic development for memory disorders. Our integrated approach demonstrates how cross-species signaling pathway analysis and carefully aligned behavioral paradigms can improve the predictive validity of preclinical models in drug discovery pipelines.
Episodic memory enables the recollection of past experiences, including specific items and their contextual details ("what," "where," and "when"). This capacity deteriorates in numerous neuropsychiatric and neurodegenerative disorders, creating an urgent need for effective therapeutics. Rodent models are indispensable in this pursuit, yet 90% of neuropsychiatric drugs that show efficacy in animal models fail in human clinical trials, often due to species-specific physiological differences and inadequate behavioral translation [91].
The hippocampal formation plays a central role in item-in-context memory across species, though its operational principles manifest differently. Research indicates the hippocampus balances two seemingly opposing functions: pattern separation (differentiating similar experiences) and pattern completion (generalizing across shared elements) [92]. This balance is crucial for adaptive memory function and represents a key target for therapeutic intervention.
This Application Note provides a comprehensive framework for cross-species investigation of item-in-context memory, with detailed protocols designed to maximize translational validity for researchers and drug development professionals.
Theoretical Framework: Item-in-Context Memory Across Species
Neural Mechanisms of Item-in-Context Binding
Item-in-context memory relies on a distributed network with the hippocampus as a central hub. The prevailing model suggests:
- Perirhinal cortex processes item information ("what")
- Parahippocampal cortex processes contextual information ("where")
- Hippocampus binds item and context into unified representations [92]
Human fMRI studies reveal that hippocampal activity patterns discriminate between events that share either item or context information, but generalize across events that share similar item-context associations [92]. This neural organization supports both highly specific memories and appropriate generalization across similar experiences.
Cross-Species Comparability
While fundamental memory processes are conserved across mammals, significant species differences exist:
- Rodents excel at spatial and contextual memory tasks
- Humans demonstrate more complex temporal and relational binding
- Primate models offer closer physiological similarity but present practical challenges [91]
Recent evidence suggests a third process, "context familiarity," exists alongside the established duality of recollection and item familiarity, representing a qualitatively distinct memory process with unique electrophysiological signatures [93]. This tri-component model (item familiarity, context familiarity, and recollection) offers a more nuanced framework for investigating memory across species.
Quantitative Cross-Species Comparison Tables
Table 1: Comparison of Species Characteristics for Memory Research
| Parameter | Mouse Models | Rat Models | Non-Human Primates | Human Applications |
|---|---|---|---|---|
| Phylogenetic Similarity to Humans | Moderate | Moderate | High | N/A |
| Research Practicalities | Short reproductive cycle, economical maintenance | Short reproductive cycle, economical maintenance | High costs, extended growth cycles, ethical considerations | Direct clinical relevance |
| Physiological Relevance | Significant differences in cardiovascular, neurophysiological systems | Closer than mice but still significant differences | High similarity in neurophysiology and cardiovascular systems | Gold standard for clinical translation |
| Item-in-Context Memory Paradigms | Well-established behavioral tasks | Extensive behavioral literature available | Limited but growing task development | Direct testing with sophisticated paradigms |
| Drug Development Predictive Value | Limited for vascular aging drugs [91] | Limited for vascular aging drugs [91] | Higher predictive value for vascular aging drugs [91] | Clinical trial outcomes |
| Key Advantages | Genetic manipulability, rapid testing | Well-characterized neurobehavioral profiles | Closest neurobiological similarity | Direct clinical relevance |
| Key Limitations | Significant physiological differences from humans | Significant physiological differences from humans | High costs, ethical considerations | Limited experimental access |
Table 2: Cross-Species Decision-Making Parameters in Perceptual Tasks
| Parameter | Mice | Rats | Humans |
|---|---|---|---|
| Average Accuracy | Lowest accuracy among species | Intermediate accuracy | Highest accuracy |
| Response Time | Fastest responses | Intermediate response times | Slowest responses |
| Decision Threshold (DDM) | Lowest decision bounds | Intermediate decision bounds | Highest decision bounds |
| Primary Behavioral Strategy | Switch between evidence accumulation and other strategies | Optimize for reward rate | Prioritize accuracy over speed |
| Internal Time Pressure | Present (collapsing bounds) | Present (collapsing bounds) | Less evidence of time pressure |
| Inter-Subject Variability | High animal-to-animal variability | Moderate variability | Lower variability |
Experimental Protocols
Cross-Species Signaling Pathway Analysis Protocol
This protocol validates animal models for drug screening by analyzing conserved and divergent molecular pathways across species [91].
Materials:
- RNA-sequencing data (bulk and single-cell) from target tissues
- Species: Rat (Rattus norvegicus), monkey (Macaca fascicularis), human (Homo sapiens)
- Software: OrthoVenn3 for phylogenetic analysis, Seurat V4 for scRNA-seq analysis, GSEA 4.3.2 for pathway enrichment
Procedure:
- Data Collection and Processing
- Obtain bulk and single-cell RNA-sequencing data from blood vessels or relevant tissues across species
- Process data using consistent normalization pipelines
- For scRNA-seq: Reduce dimensionality using PCA and cluster cells based on gene expression similarity
Phylogenetic Relationship Analysis
- Use OrthoVenn3 to infer evolutionary relationships between species
- Identify orthologous genes with conserved expression patterns
Differential Expression Analysis
- Identify genes with consistent/differential expression patterns across species
- Focus on age-related changes relevant to the disease process
Pathway Enrichment Analysis
- Perform Gene Set Enrichment Analysis (GSEA) using pre-ranked gene lists
- Calculate Normalized Enrichment Scores (NES) to determine pathway activation (+) or inhibition (-)
- Compare pathway trends across species using NES values
Protein-Protein Interaction Network Analysis
- Upload differentially expressed genes to STRING database
- Construct PPI networks and identify key nodes using betweenness centrality algorithm in Cytoscape
Model Validation
- Select pathways showing consistent trends across species for targeted drug testing
- Compare pharmacological predictions with known drug effects in animal/clinical experiments
- Use inconsistent pathways to explain drug failures or adverse effects
Applications:
- Validation of animal models for specific disease pathways
- Prediction of drug efficacy and potential adverse effects
- Identification of conserved therapeutic targets
Item-in-Context Memory Task Protocol
This protocol assesses integrated memory for items and their contextual associations across species-appropriate implementations.
Materials:
- Rodents: Operant chambers with nose ports, visual stimulus panels, reward delivery system
- Humans: Computer-based task with visual stimuli, response input device
- Stimuli: Category-specific images (objects, faces), contextual cues (locations, backgrounds)
Procedure:
- Stimulus Preparation
- Select 28 categories of visual objects with 12 unique exemplars per category
- Assign 8 objects per category as targets, 4 as foils
- Designate 4 distinct context locations (e.g., park, beach, restaurant, café)
Context Personalization (Human Subjects)
- Instruct participants to select specific personal locations for each context cue
- Ensure participants can vividly visualize these venues
Encoding Phase
- Present items in mini-blocks of 2-5 items per context
- Show context cue for 3 seconds before each mini-block
- Present each item for 2 seconds with instructional query
- For rodents: Use olfactory or spatial contexts instead of verbal cues
- Counterbalance item-context pairings across subjects
Retrieval Phase
- Present items (targets and foils) in random order
- Require context judgment for each recognized item
- Record accuracy and response time for item and context memory
Data Analysis
- Calculate item recognition accuracy (hits vs. false alarms)
- Calculate source memory accuracy (correct context attribution)
- Use signal detection theory to compute d' for item and context memory
- Analyze reaction time differences between conditions
Variations:
- Temporal Binding Protocol: Assess memory for serial order within and across event boundaries [22]
- Free Recall Protocol: Test uncued retrieval of item sequences [34]
Visualization of Cross-Species Validation Framework
Diagram 1: Integrated Framework for Cross-Species Validation of Memory Mechanisms. This workflow integrates molecular and behavioral analysis tiers to identify conserved and divergent pathways for predictive model selection.
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Essential Research Reagents and Solutions for Item-in-Context Memory Research
| Tool/Reagent | Function | Species Applicability | Key Considerations |
|---|---|---|---|
| Single-cell RNA-sequencing | Cell-type specific transcriptomic profiling | All species | Enables identification of conserved cell populations and signaling pathways [91] |
| fMRI with multivariate pattern analysis | Non-invasive measurement of distributed neural representations | Humans, NHPs | Detects pattern separation and completion in hippocampal subfields [92] |
| Operant Chambers with Nose Ports | Automated behavioral testing for rodents | Rodents | Enable high-throughput testing of item-context associations with precise stimulus control |
| Drift Diffusion Modeling (DDM) | Computational modeling of decision processes | All species | Quantifies decision thresholds, drift rates, and non-decision time across species [94] |
| OrthoVenn3 Software | Phylogenetic relationship analysis | Cross-species | Identifies orthologous genes and evolutionary relationships [91] |
| STRING Database | Protein-protein interaction network analysis | Cross-species | Maps interactions between conserved gene products across species [91] |
| Event-Related Potentials (ERPs) | Physiological measurement of memory processes | Humans, NHPs | Differentiates item familiarity, context familiarity, and recollection [93] |
| Pulse-Based Evidence Accumulation Task | Perceptual decision-making assessment | All species | Synchronized framework enables direct cross-species comparison [94] |
Applications in Drug Development
Target Validation and Prioritization
The cross-species signaling pathway analysis framework enables target prioritization based on evolutionary conservation. Drugs targeting pathways with consistent expression trends across species demonstrate higher translational success, while those targeting divergent pathways frequently exhibit adverse effects or efficacy loss [91].
Model Selection for Specific Pathways
Different animal models show varying predictive validity depending on the targeted pathway:
- Rodent models are suitable for highly conserved neural pathways
- Non-human primates are essential for complex cognitive functions with cortical dependencies
- Cross-species validation is critical for pathways showing species-specific regulation
Cognitive Biomarker Development
Synchronized behavioral frameworks enable development of cross-species cognitive biomarkers that can track treatment effects from preclinical to clinical stages. The pulse-based evidence accumulation task demonstrates how quantitative comparison of decision parameters can bridge species gaps in perceptual and cognitive assessment [94].
Cross-species validation of rodent models for item-in-context memory requires integrated molecular, behavioral, and computational approaches. By implementing the detailed protocols and frameworks presented in this Application Note, researchers can significantly improve the predictive validity of preclinical models, ultimately accelerating the development of effective therapeutics for memory disorders. The future of translational memory research lies in standardized, cross-species validated approaches that acknowledge both conserved principles and species-specific adaptations in neural function.
Conclusion
Temporal binding tasks provide a powerful, quantifiable framework for investigating the dynamic processes underlying episodic memory formation and distortion. The research synthesized demonstrates that Temporal Binding Windows are malleable neurocognitive constructs influenced by pharmacological interventions, clinical conditions, and perceptual training. Key takeaways include: (1) Psychoactive drugs exert stage-specific effects on emotional episodic memory, with important implications for both addiction mechanisms and therapeutic development; (2) TBW alterations serve as sensitive markers for neuropsychiatric conditions like OCD and prodromal neurodegenerative states; (3) Computational models integrating multiple timescales of synaptic plasticity successfully predict behavioral outcomes in episodic memory tasks. Future directions should focus on developing TBW-based biomarkers for early disease detection, designing targeted pharmacological interventions that selectively modulate emotional memory processes, and exploring non-pharmacological TBW training as adjunctive therapy for conditions characterized by maladaptive memory patterns. These advances position temporal binding research at the forefront of innovative memory modification therapies and precision medicine approaches in psychiatry and neurology.
References
- 1. The Construct of the Multisensory Temporal Binding Window ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4326640/]
- 2. Perceptual Training Narrows the Temporal Window of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2771316/]
- 3. Perceived Simultaneity and Temporal Order of Audiovisual ... [https://www.frontiersin.org/journals/human-neuroscience/articles/10.3389/fnhum.2018.00139/full]
- 4. Negative emotion reduces the temporal compression of ... [https://pubmed.ncbi.nlm.nih.gov/40340765/]
- 5. Temporal integration as an adaptive process in visual ... [https://www.sciencedirect.com/science/article/pii/S0149763425000417]
- 6. Segregation of complex acoustic scenes based on ... [https://elifesciences.org/articles/00699]
- 7. Understanding Episodic Memory Networks - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC12314160/]
- 8. Hippocampal-cortical interactions in the consolidation of ... [https://www.nature.com/articles/s41467-025-64264-7]
- 9. Representation of memories in the cortical-hippocampal system [https://pmc.ncbi.nlm.nih.gov/articles/PMC4930430/]
- 10. The Hippocampus Supports High-Resolution Binding in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3773061/]
- 11. Hippocampal neurons code individual episodic memories ... [https://www.nature.com/articles/s41562-023-01706-6]
- 12. Increased cortical plasticity leads to memory interference ... [https://elifesciences.org/articles/84911]
- 13. Hippocampal Activity Patterns Carry Information about ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3984944/]
- 14. The role of working memory in encoding the temporal ... [https://pubmed.ncbi.nlm.nih.gov/40991194/]
- 15. The semantic structure of events consistently influences ... [https://www.nature.com/articles/s41598-025-26142-6]
- 16. Temporal binding of interval markers | Scientific Reports [https://www.nature.com/articles/srep38806]
- 17. Time for Action: Verbal Action Cues Influence Temporal ... [https://www.frontiersin.org/journals/psychology/articles/10.3389/fpsyg.2020.00160/full]
- 18. The spatiotemporal organization of episodic memory and ... [https://www.nature.com/articles/s41598-019-53823-w]
- 19. Episodic boundaries affect neural features of ... [https://elifesciences.org/reviewed-preprints/90576v1]
- 20. A position coding model that accounts for the effects of ... [https://pubmed.ncbi.nlm.nih.gov/39818105/]
- 21. Examining the development of memory for temporal ... [https://www.sciencedirect.com/science/article/pii/S1878929321000232]
- 22. Temporal binding within and across events - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC5018468/]
- 23. Short-term plasticity influences episodic memory recall [https://www.nature.com/articles/s41598-025-12611-5]
- 24. How the New Interacts With the Old? Hippocampal Processing ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12547841/]
- 25. Is the temporal binding effect in the Libet clock-task based ... [https://www.sciencedirect.com/science/article/pii/S1053810025001023]
- 26. Bayesian Temporal Prediction: A Robust Algorithm for Real ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12395933/]
- 27. A contextual binding theory of episodic memory - PMC - NIH [https://pmc.ncbi.nlm.nih.gov/articles/PMC7233541/]
- 28. Effects of multisensory simultaneity judgment training on ... [https://www.nature.com/articles/s41598-025-96121-4]
- 29. Perceiving Audiovisual Synchrony: A Quantitative ... [https://www.sciencedirect.com/science/article/abs/pii/S0149763425004506]
- 30. Correlation detection as a stimulus computable account for ... [https://elifesciences.org/articles/106122]
- 31. Perceptual training of audiovisual simultaneity judgments ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12238671/]
- 32. A Quantitative Synthesis of Simultaneity and Temporal ... [https://pubmed.ncbi.nlm.nih.gov/41167444/]
- 33. Episodic Memory [https://thedecisionlab.com/reference-guide/psychology/episodic-memory]
- 34. Neuroscientist's Behavioral Toolbox for Studying Episodic- ... [https://www.eneuro.org/content/11/8/ENEURO.0073-24.2024]
- 35. Memory-Guided Saccades in Subacute and Chronic Stroke [https://www.mdpi.com/2227-9059/12/8/1678]
- 36. Neural Dynamics of Relational Memory Retrieval Across Eye ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12612354/]
- 37. Saccades track visual associative memory processes with ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12204191/]
- 38. The acute effects of psychoactive drugs on emotional episodic ... [https://pubmed.ncbi.nlm.nih.gov/37085021/]
- 39. The Development of Episodic Memory: Items, Contexts, ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9295207/]
- 40. Αlpha 5 subunit-containing GABAA receptors in temporal lobe ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7811756/]
- 41. Episodic memory and inhibitory control in people who inject ... [https://bmcpsychiatry.biomedcentral.com/articles/10.1186/s12888-025-07223-2]
- 42. Drug-Induced Cognitive Impairment - PMC - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC11903592/]
- 43. GABA-A receptor modulating steroids in acute and chronic ... [https://www.sciencedirect.com/science/article/pii/S235228951930058X]
- 44. Largest Study Ever Done on Cannabis and Brain Function ... [https://news.cuanschutz.edu/news-stories/largest-study-ever-done-on-cannabis-and-brain-function-finds-impact-on-working-memory]
- 45. Brain Function Outcomes of Recent and Lifetime Cannabis Use [https://pubmed.ncbi.nlm.nih.gov/39874032/]
- 46. Cannabis and Nicotine/Tobacco Co-Use and its ... [https://www.sciencedirect.com/science/article/pii/S0149763425004816?dgcid=rss_sd_all]
- 47. The effect of biperiden on episodic memory: Testing ... [https://www.sciencedirect.com/science/article/pii/S1074742725000462]
- 48. Dopaminergic processes predict temporal distortions in event ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12132335/]
- 49. Temporal Modulation of Spike-Timing-Dependent Plasticity [https://pmc.ncbi.nlm.nih.gov/articles/PMC3059714/]
- 50. A simple model for Behavioral Time Scale Synaptic ... [https://www.nature.com/articles/s41467-024-55563-6]
- 51. Estimating Brain Activity with High Spatial and Temporal ... [https://arxiv.org/html/2510.09415v1]
- 52. Frontiers | Epigenetic mechanisms of plasticity and resistance in glioblastoma: therapeutic targets and implications [https://www.frontiersin.org/journals/epigenetics-and-epigenomics/articles/10.3389/freae.2025.1519449/full]
- 53. Methodological Factors Involved in the Study of Temporal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7266961/]
- 54. Confident failures: Lapses of working memory reveal a ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5520639/]
- 55. The role of working memory capacity in the temporal compression of episodic memories: An individual differences approach - PubMed [https://pubmed.ncbi.nlm.nih.gov/38913729/]
- 56. Temporal binding task [https://bio-protocol.org/exchange/minidetail?id=18633486&type=30]
- 57. Focusing attention in working and long-term memory ... [https://www.nature.com/articles/s41467-025-59359-0]
- 58. The acute effects of psychoactive drugs on emotional ... [https://www.sciencedirect.com/science/article/abs/pii/S0149763423001574]
- 59. The order of multisensory associative sequences is ... [https://www.nature.com/articles/s41598-025-02553-3]
- 60. The Mechanisms and Boundary Conditions of Drug Memory ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8377231/]
- 61. Offline consolidation mechanisms of the retrieval practice ... [https://www.nature.com/articles/s41539-025-00349-8]
- 62. Attention and feature binding in the temporal domain - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC11680606/]
- 63. The intrinsic time tracker: temporal context is embedded in ... [https://www.nature.com/articles/s41467-025-63633-6]
- 64. Locating Temporal Functional Dynamics of Visual Short ... [https://www.nature.com/articles/srep42013]
- 65. A temporal comparison of retro-cue effects in visual ... [https://link.springer.com/article/10.3758/s13421-025-01789-8]
- 66. Optimizing electrical stimulation parameters to enhance visual ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12271465/]
- 67. Investigation of the optimal time interval between task ... [https://www.sciencedirect.com/science/article/pii/S0167876022001209]
- 68. Dynamic emotional states shape the episodic structure of ... [https://www.nature.com/articles/s41467-023-42241-2]
- 69. The developmental profile of temporal binding [https://pmc.ncbi.nlm.nih.gov/articles/PMC7534204/]
- 70. The fusion point of temporal binding: Promises and perils ... [https://www.sciencedirect.com/science/article/pii/S0010028524000331]
- 71. Ten simple rules for the computational modeling of ... [https://elifesciences.org/articles/49547]
- 72. Color contrast - Accessibility - MDN Web Docs - Mozilla [https://developer.mozilla.org/en-US/docs/Web/Accessibility/Guides/Understanding_WCAG/Perceivable/Color_contrast]
- 73. Elements must meet minimum color contrast ratio thresholds [https://dequeuniversity.com/rules/axe/4.8/color-contrast]
- 74. Understanding episodic memory dynamics: Retrieval and ... [https://www.sciencedirect.com/science/article/pii/S1053811925001727]
- 75. Presentation of Quantitative Data [https://ihatepsm.com/blog/presentation-quantitative-data]
- 76. Presenting Quantitative Data Graphically [https://courses.lumenlearning.com/waymakermath4libarts/chapter/presenting-quantitative-data-graphically/]
- 77. Exploring Visuo-Tactile Temporal Binding Window ... [https://pubmed.ncbi.nlm.nih.gov/40895833/]
- 78. Exploring Visuo-Tactile Temporal Binding Window ... [https://www.sciencedirect.com/science/article/pii/S2667174325001193]
- 79. Reduced body cell mass and functions in lower extremities ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10435546/]
- 80. Alterations in Striatal Circuits Underlying Addiction-Like ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5523013/]
- 81. Temporal binding during deliberate rule breaking [https://www.sciencedirect.com/science/article/pii/S1053810025000443]
- 82. Episodic memory development: Bridging animal and ... [https://www.sciencedirect.com/science/article/pii/S0896627324000461]
- 83. From Correlation to Causation: Understanding Episodic ... [https://link.springer.com/article/10.1007/s12264-025-01407-2]
- 84. Across time and space: spatial-temporal binding under stress [https://pmc.ncbi.nlm.nih.gov/articles/PMC6859825/]
- 85. Assessing Saccadic Eye Movements With Head-Mounted ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7527608/]
- 86. Saccadic Eye Movement in Mild Cognitive Impairment and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9090874/]
- 87. Saccadic eye movements differentiate functional cognitive ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12417601/]
- 88. Eye Tracking Smooth Pursuit and Saccades [https://www.sr-research.com/case-studies/eye-tracking-smooth-pursuit-and-saccades/]
- 89. A New Algorithm for Identifying Saccadic Movements in ... [https://iotlenses.com/blog/algorithm-to-identify-saccadic-movements-eye-tracking-studies.html]
- 90. Computational models of working memory: putting long- ... [https://www.sciencedirect.com/science/article/abs/pii/S1364661305002731]
- 91. Cross‐species signaling pathways analysis inspire animal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11164676/]
- 92. The hippocampus generalizes across memories that share ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7217712/]
- 93. Context familiarity is a third kind of episodic memory ... [https://www.sciencedirect.com/science/article/pii/S2589004224026646]
- 94. A cross-species framework for investigating perceptual ... [https://elifesciences.org/reviewed-preprints/105116]